")
Back to Journals » Clinical Interventions in Aging » Volume 20
Adipose Factor ANGPTL4: Its Role in Aging Mechanisms and Associated Diseases
Authors Wang X, Chang HC, Gu X , Zhang Y, Bao Z
Received 11 February 2025
Accepted for publication 4 June 2025
Published 28 June 2025 Volume 2025:20 Pages 911—929
DOI https://doi.org/10.2147/CIA.S522049
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Maddalena Illario
Xiaojun Wang,1,2,* Hung-chen Chang,1,2,* Xuchao Gu,1,2,* Yuxin Zhang,1,2 Zhijun Bao1– 3
1Shanghai Key Laboratory of Clinical Geriatric Medicine, Huadong Hospital Affiliated to Fudan University, Shanghai, 200040, People’s Republic of China; 2Shanghai Institute of Geriatric Medicine, Huadong Hospital Affiliated to Fudan University, Shanghai, 200040, People’s Republic of China; 3Department of Gerontology, Huadong Hospital Affiliated to Fudan University, Shanghai, 200040, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhijun Bao; Yuxin Zhang, Shanghai Key Laboratory of Clinical Geriatric Medicine, Huadong Hospital Affiliated to Fudan University, Shanghai, 200040, People’s Republic of China, Email [email protected]; [email protected]
Abstract: The angiopoietin-like protein 4 (ANGPTL4), also known as fasting-induced adipose factor, is a secreted glycoprotein that belongs to the ANGPTL protein family. Due to its expression in various cell types and tissues and its interactions with other proteins, ANGPTL4 plays diverse roles within its family, exhibiting a wider range of molecular functions. For instance, ANGPTL4 is intricately involved in modulating central energy metabolism and enhancing exercise endurance, while also acting as a pivotal mediator in the interaction between gut microbiota and host lipid metabolism. Moreover, the expression of ANGPTL4 is directly controlled by aging-related signaling pathways. Its excessive activation accelerates the aging process by triggering mechanisms like heightened oxidative stress, epithelial-mesenchymal transition (EMT) and fibrosis, abnormal lipid accumulation, and cellular arrest, thereby advancing the development of age-related diseases. Given the pivotal roles of ANGPTL4 and its associated molecules in organ fibrosis and cancer advancement, targeting ANGPTL4 emerges as a promising therapeutic approach. However, the intricate and sometimes conflicting functions of the two cleavage fragments of ANGPTL4, namely N-terminal fragment (nANGPTL4) and C-terminal fragment (cANGPTL4), in different chronic diseases—exerting inhibitory or stimulatory effects depending on the disease stage—have posed challenges to the progress of ANGPTL4 antibody therapy. This review provides an overview of the biological mechanisms of ANGPTL4, its dual impact on fibrosis and tumorigenesis, and highlights its recent advancements as a potential biomarker in age-related diseases and inflammation-related conditions. ANGPTL4 is a high-potential but complex target, requiring mechanism-driven strategies for safe clinical translation.
Keywords: ANGPTL4, adipose factor, age-related diseases, mechanism
Introduction
In the year 2000, multiple research groups initially characterized fasting-induced adipose factor (FIAF), also known as Angiopoietin-like 4 (ANGPTL4). ANGPTL4 is a member of the ANGPTL family, a glycoprotein comprising 406 amino acids with a molecular mass ranging from approximately 45 to 65 kDa, and it is encoded by the 19p13.2 locus.1 Structurally, ANGPTL4 contains an N-terminal coiled-coil domain (CCD) with α-helical oligomerization capability, and a C-terminal fibrinogen-like domain (FLD) that can be subdivided into A, B, and P subdomains. Experimental evidence has validated the diverse biological functions associated with these two structural domains (CCD and FLD).2 The protein structure prediction data referenced in this study are sourced from the AlphaFold database3,4 (version 2022–11-01, Monomer v2.0), and the data are used under the CC-BY-4.0 license (Figure 1).
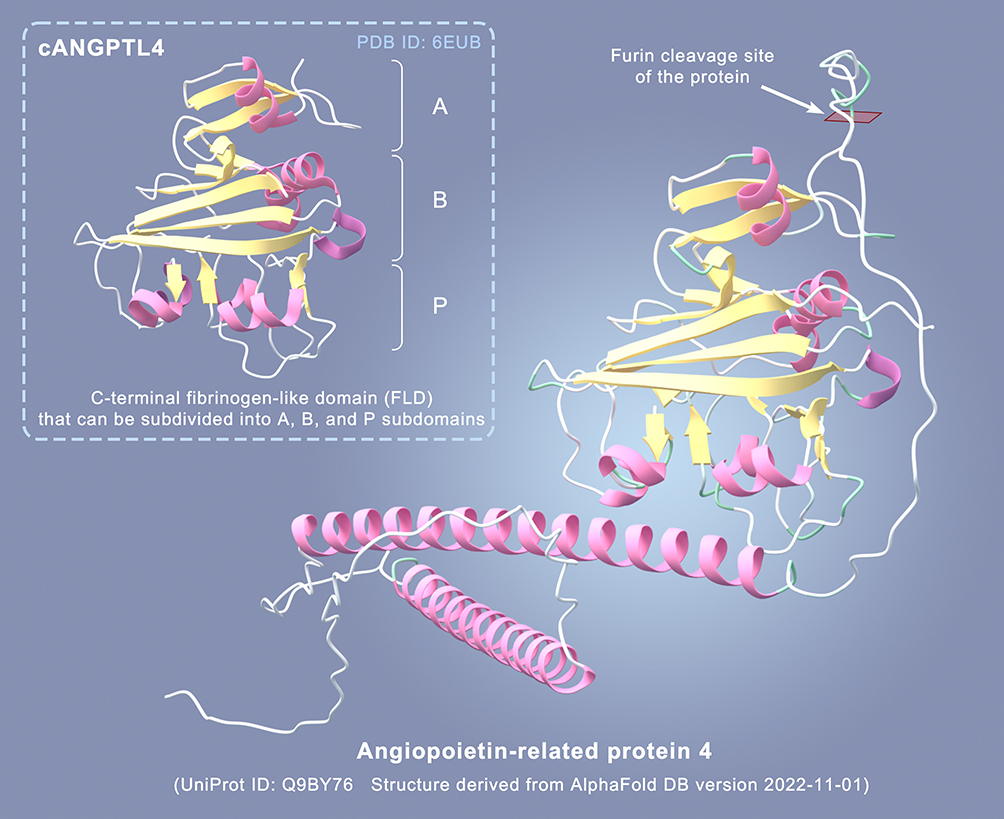 |
Figure 1 Different Structural Domains and Cleavage Sites of ANGPTL4. |
The secreted protein ANGPTL4 is primarily expressed in adipose tissue and the liver.5 The cleavage forms of ANGPTL4 exhibit tissue-specificity, with the liver producing the N-terminal fragment N-ANGPTL4 (nANGPTL4) and adipose tissue generating the full-length ANGPTL4 protein. The full-length ANGPTL4 protein is 52 kDa in size, with cleavage occurring after the 164th amino acid. The nANGPTL4 is 15 kDa, and the C-terminal fragment C-ANGPTL4 (cANGPTL4) is 37 kDa. This differential cleavage may contribute to varied physiological functions and regulatory effects in different tissues.6 cANGPTL4 triggers fat breakdown by elevating cAMP levels in adipocytes. Adenoviral overexpression of cAngptl4 boosts oxygen consumption, fat utilization, and the expression of brown fat thermogenesis genes (Ucp1 and Ppargc1a) in subcutaneous white adipose tissue of mice, resulting in reduced body weight in high-fat diet-fed mice without altering lipoprotein lipase (LPL) activity.7 Hepatocytes are capable of both expressing and cleaving ANGPTL4. Hepatocyte-expressed Furin, PC5/6, PACE, and PC7 proteases can recognize the proprotein convertase (PC) consensus cleavage sequence RXKR (with species-specific amino acid arrangement at positions 180 to 183 of the ANGPTL4 protein, RRKR for humans, RGKR for rats and mice), leading to a significant reduction in the levels of full-length ANGPTL4 and cANGPTL4.1 In vitro studies have demonstrated that a truncated fragment of the CCD domain of nANGPTL4 can inhibit the formation and activity of RAW264.7 osteoclasts, while also mildly promoting osteoblast proliferation, without impacting osteoblast differentiation in vitro. Conversely, the full-length and C-terminal FLD domains do not display these particular functional properties.8
ANGPTL4, identified as a LPL inhibitor, serves as an endogenous, non-competitive regulator involved in the modulation of lipid deposition and energy homeostasis.9 Both the full-length and N-terminal forms of ANGPTL4 exhibit inhibitory effects on LPL, while the C-terminal fragment shows no impact on LPL activity. In vitro investigations have demonstrated that the cleavage products of ANGPTL4 possess heightened potency as LPL inhibitors.10 The inhibition of LPL is facilitated by a specific short amino acid sequence in close proximity to the N-terminus of ANGPTL4. Molecular models suggest that the nANGPTL4 triggers the transition of active LPL dimers into inactive monomers.11 ANGPTL4 induces the inactivation of LPL monomers by triggering the irreversible unfolding of the LPLα/β hydrolase domain. This unfolding process is initiated when ANGPTL4 binds to sequences located near the LPL catalytic site, specifically within the β2, β3-α3, and lid regions. The binding of ANGPTL4 induces conformational changes at the β3-α3 region, which subsequently propagate to β5 and β4-α4, ultimately resulting in the irreversible unfolding of the LPL catalytic pocket region.12 Moreover, ANGPTL4 can deactivate LPL either in the endothelial interstitium or directly at the cell surface before exerting systemic regulatory effects.13 Additionally, ANGPTL4 exhibits the capability to enhance the cleavage of LPL by PC, a function that remains unaffected by its own cleavage status.14
The Distinctiveness of ANGPTL4
Different from other family members, the expression of ANGPTL4 is influenced by various factors such as diet, hormone levels, and exercise.15,16 Research has confirmed that exercise-induced ANGPTL4 is secreted by the liver and is regulated by the human glucagon-cAMP-PKA pathway.17 Furthermore, studies have revealed a new role of skeletal muscle ANGPTL4 in exercise physiology.18 Treatment with ANGPTL4 activates AMP-activated protein kinase (AMPK) signaling in skeletal muscle cells. Conversely, the absence of ANGPTL4 diminishes exercise-induced AMPK activation in skeletal muscle, leading to reduced exercise endurance in ANGPTL4-deficient mice. Interestingly, ANGPTL4 is also influenced by temperature regulation. Studies have demonstrated that ANGPTL4 plays a crucial role in regulating plasma lipid distribution during prolonged cold exposure. ANGPTL4 can guide fatty acids from triglyceride-rich lipoproteins in circulation to brown adipose tissue during cold exposure.19
ANGPTL4 also appears to play a significant role in the central regulation of energy metabolism.20 An in vivo study revealed that the targeted deletion of ANGPTL4 increased the availability of fatty acids in astrocytes, thereby preventing diet-induced obesity.21 In another in vitro study, primary hippocampal neurons responded to saturated fatty acids by upregulating the expression of ANGPTL4 and pro-inflammatory factors, suggesting a potential involvement of ANGPTL4 in neuronal damage associated with metabolic diseases.22 More Interestingly, central ANGPTL4 does not directly regulate LPL activity but seems to participate in metabolic communication between glial cells and neurons.23 Moreover, ANGPTL4 acts as a natural inhibitor of intestinal lipase activity.24 Research indicates that ANGPTL4 can be synthesized by enteroendocrine cells in the human intestinal tract, with its expression potentially modulated by short-chain fatty acids and bile acids.25 ANGPTL4 may also enhance bile acid absorption during tauroursodeoxycholic acid supplementation through mechanisms dependent on the intestinal microbiota.26 Clinical studies have revealed interactions between microbial baiCD genes and host ANGPTL4 genes in the non-inflammatory intestinal mucosa of patients with inflammatory bowel disease.27
Emerging evidence suggests a potential involvement of ANGPTL4 in the aging process.28,29 The Wnt/β-catenin signaling pathway diminishes with age,30,31 while ANGPTL4 acts as an antagonist of the Wnt signal by promoting LRP6 turnover.32 An in vitro experiment confirmed that ANGPTL4 can inhibit C2C12 differentiation by suppressing the Wnt/β-catenin signaling pathway.33 Moreover, with advancing age, there is an increased accumulation of lipid droplets around the nuclear membrane.34 Our preliminary investigations have indicated that the upregulation of the adipokine ANGPTL4 in proximal tubule cells is associated with lipid droplet accumulation and accelerated aging.35 More importantly, a study indicated that the overexpression of ANGPTL4 exacerbated the aging phenotype in cardiomyocytes induced by high glucose and palmitic acid, leading to increased markers of aging, while SGLT2 was shown to mitigate this effect.36 The research suggests that targeting ANGPTL4 (such as with SGLT2 inhibitors) may alleviate age-related cardiac dysfunction. Unlike ANGPTL3 and ANGPTL8 (mainly liver-derived), ANGPTL4 is broadly induced by aging-associated stressors (eg, hypoxia, oxidative stress) in adipose tissue, endothelium, and senescent cells.37,38 This suggests its potential role as a stress-responsive mediator in aging. Although few studies directly compare ANGPTL members in aging models, existing data suggest ANGPTL4’s broader involvement in aging-related pathways.
Presently, there is a dearth of comprehensive reviews addressing the role of ANGPTL4 in aging-related mechanisms. Aging is primarily driven by conservative features, including genomic instability, mitochondrial dysfunction, and chronic inflammation.39 Notably, processes regulated by ANGPTL4, such as oxidative stress accumulation and epithelial-mesenchymal transition (EMT), directly intersect with these features.40,41 EMT can promote the development of these aging hallmarks, hastening cellular functional decline and tissue homeostasis imbalance.42 This manuscript predominantly focuses on elucidating the multifaceted functions of ANGPTL4 in aging processes, encompassing angiogenesis, EMT, oxidative stress modulation, and the senescence-associated secretory phenotype (SASP) secretion. Furthermore, it explores the prospective utility of ANGPTL4 as a biomarker in age-related diseases.
The Potential Association Between ANGPTL4 and Age-Related Mechanisms
Angiogenesis
ANGPTL4 plays a crucial role in maintaining vascular homeostasis, age-related vascular dysfunction, and tumor angiogenesis. A groundbreaking study has discovered that ANGPTL4 improves age-related vascular dysfunction. This study proposes that elevating ANGPTL4 levels in specific aging tissues, such as the vascular system or brain endothelium, could offer protective and potentially therapeutic benefits.43 Research suggests that hypoxia stimulates brain endothelial cell migration and vascular malformation remodeling through astrocyte-derived ANGPTL4. Furthermore, in the oxygen-induced retinopathy model, ANGPTL4 deficiency markedly suppresses hypoxia-driven pathological neovascularization, providing further evidence of the crucial role of ANGPTL4 in abnormal vascular proliferation.44,45 Additionally, Qiu et al have validated that ANGPTL4 enhances post-stroke angiogenesis and neurogenesis, reduces neuronal death, and suppresses inflammation by upregulating AKT phosphorylation.46 Another study proposes that mechanical loading can trigger tenocytes to increase the expression of ANGPTL4 through the TGF-β/HIF-1α pathway, potentially modulating local vascular responses and impacting the tendon repair process.47 In terms of vascular homeostasis, ANGPTL4 serves as a crucial regulator of vascular integrity under both developmental and hypoxia-induced pathological conditions. By modulating integrin αvβ3 binding, VEGFR2-Src kinase signaling, and endothelial junction stability, ANGPTL4 can mitigate hypoxia-driven vascular permeability.48 Studies have further shown that mice with endothelial cell-specific deletion of ANGPTL4 exhibit diminished pathological neovascularization and improved vascular stability.49
In addition to its roles in aging and vascular protection, ANGPTL4 exhibits distinct functions in the tumor microenvironment. Research has elucidated that ANGPTL4 can facilitate ovarian cancer progression via the JAK signaling pathway and promote angiogenesis within the tumor microenvironment through its interaction with ESM1.50 Experimental data reveals that genetic deletion or antibody-mediated neutralization of ANGPTL4 leads to a substantial reduction in obesity-induced angiogenesis and tumor growth.51 These observations underscore the critical role of ANGPTL4 in modulating tumor angiogenesis.
Intriguingly, ANGPTL4 appears to have a dual role in vascular endothelial dysfunction. Its different protein forms - nANGPTL4 and cANGPTL4 - exhibit distinct and even contradictory functions, depending on the tissue microenvironment, protein cleavage status, and activation of downstream signaling pathways. Studies have found that C-Angptl4, rather than N-Angptl4, effectively inhibits endothelial cell proliferation, migration, and tube formation, preventing neovascularization in mice. Mechanistically, cANGPTL4 suppresses angiogenesis by inhibiting the Raf/MEK/ERK1/2 MAP kinase signaling pathway in endothelial cells.52 Preclinical studies in multiple tumor models have shown that in VEGF-A-induced vascular generation, both cANGPTL4 and nANGPTL4 have contrasting effects. Specifically, cANGPTL4 stimulation alone results in a mild vascular budding response, while a combination of low-dose VEGF-A with full-length ANGPTL4 or cANGPTL4 leads to a strong vascular generation response. In contrast, nANGPTL4 does not affect vascular budding and effectively inhibits VEGF-A-induced vascular generation.53 Another study demonstrated that cANGPTL4 binds and activates integrin α5β1-mediated Rac1/PAK signaling, contributing to endothelial disruption.54 This characteristic makes ANGPTL4 a potential target for treating vascular diseases, but precise regulation of specific segments is required.
Epithelial-Mesenchymal Transition (EMT)
The EMT is a well-established biological process implicated in pathological physiological phenomena such as tissue fibrosis, organ aging, and functional decline.55 During EMT, damaged epithelial cells can play a significant role as a source of fibroblasts.56 Recent research from our team has unveiled that aging-related renal tubular epithelial cells exhibit an upregulation in ANGPTL4 protein levels, potentially further promoting renal fibrosis progression.35 Moreover, recent investigations have identified selective expression of ANGPTL4 in the fibroblast area of idiopathic pulmonary fibrosis lung tissue, concomitant with an increase in the expression of the myofibroblast marker α-SMA.57 Another study proposes that fibroblast-secreted ANGPTL4 may participate in the potential mechanism of heart failure with preserved ejection fraction (HFpEF) cardiac vascular dysfunction, offering promise as a prospective drug target.58 Furthermore, modulation of the ANGPTL4 gene has been shown to regulate the expression of CD163 and the TLR4/NF-κB signaling pathway, resulting in the suppression of Kupffer cell polarization and pro-inflammatory effects, and the inhibition of hepatic stellate cell activation and fibrosis.59 Clinical studies indicate a gradual rise in ANGPTL4 levels from acute infection to liver cirrhosis and hepatocellular carcinoma (HCC), peaking in the advanced fibrotic stage.60 A recent study discovered that targeting cANGPTL4 treatment can effectively inhibit the migration of fibroblast-like synoviocytes (FLS) and inflammation-induced osteoclastogenesis.61 Collectively, these findings indicate that ANGPTL4 could play a pivotal role as a key regulatory factor in the EMT process.
The EMT is also considered a crucial initiating stage in cancer metastasis.62 Targeted therapies directed at the EMT process show potential in impeding tumor progression.63 Recent investigations have highlighted the therapeutic potential of ANGPTL4 in inhibiting EMT and arresting cancer progression.64 Studies conducted by Gordon et al have demonstrated that ANGPTL4 promotes EMT in pancreatic ductal adenocarcinoma while regulating the expression of APOL1 and ITGB4 genes. Notably, their research revealed that elevated ANGPTL4 levels led to gemcitabine resistance in vitro and reduced survival rates in patients.65 Another study revealed that oleic acid-induced secretion of ANGPTL4 enhanced migration and invasion in head and neck squamous cell carcinoma. Depletion of ANGPTL4 in cells resulted in a significant reduction in levels of EMT markers such as vimentin, MMP-9, and fibronectin.66 Teo et al identified that the ANGPTL4/14-3-3γ signaling pathway functions through specific phosphorylation signals on target proteins to regulate cellular bioenergetics, manage energy demands, and enhance tumor EMT capability and metastasis.67 Overall, research targeting ANGPTL4 as a therapeutic intervention shows promise in developing more effective strategies for cancer treatment and fibrotic disease prevention and management. Further exploration is needed to elucidate the overall mechanism of action of nANGPTL4 and cANGPTL4 in the EMT process.
Oxidative Stress Interference
Oxidative stress, resulting from the generation of free radicals in the body, including reactive oxygen species (ROS) and reactive nitrogen species (RNS), is recognized as a significant factor in the aging process.68,69 Within this context, a complex interplay exists among ANGPTL4, free fatty acids (FFA), and ROS, collectively contributing to the regulation of lipid metabolism and energy homeostasis.70,71 Singh et al observed that the absence of ANGPTL4 in liver cells facilitated fatty acid uptake, leading to enhanced fatty acid oxidation, increased ROS production, and AMPK activation.72 Additionally, studies have demonstrated that dietary fatty acids and the stimulation of cardiac ANGPTL4 gene expression by PPARβ/δ form part of a feedback mechanism aimed at shielding the heart from lipid overload and oxidative stress induced by fatty acids.73 Conversely, the lipotoxicity associated with excessive FFA levels can trigger ROS generation, cell apoptosis, and inflammatory responses.74 Our investigations indicate that the deficiency of ANGPTL4 appears to mitigate FFA-induced ROS production in HK-2 cells.35
In cancer cells, ROS play a pivotal role in the regulation and induction of apoptosis, influencing cancer cell proliferation, survival, and resistance to treatment.75 A study has identified ANGPTL4 as a significant redox factor in cancer.76 Additionally, research has shown that depletion of ANGPTL4 leads to a notable suppression of NOX4 expression and ROS production, offering a potential target for enhancing the prognosis of colorectal cancer metastasis patients with hyperlipidemia.77 Regrettably, current literature only suggests a correlation between ANGPTL4 and oxidative stress, with limited exploration into the specific mechanisms by which ANGPTL4 modulates ROS production. In this section, the cleavage state of ANGPTL4 has not been identified, and further experimental validation is needed to determine the relative contributions of the N-terminus and C-terminus.
Senescence-Associated Secretory Phenotype (SASP)
SASP is recognized for its role in promoting chronic inflammation and triggering the aging process in normal cells.78 ANGPTL4 is implicated in the context of chronic inflammation. In a clinical cohort study conducted by our research team, it was observed that in elderly frail patients, the levels of the SASP factor TNF-α were significantly and positively correlated with serum levels of ANGPTL4.79 Recent research suggests that the knockout of ANGPTL4 notably diminishes antigen-induced cell senescence promotion and reduces the expression levels of intracellular senescence markers such as IL-6, P16, and P21.80 Other studies have shown that aging zebrafish display heightened anxiety and increased central nervous system expression of ANGPTL4.81 Furthermore, augmented phosphorylation of ERK1/2 and increased expression of ANGPTL4 and the SASP factor IL-8 genes were observed in aging human skin fibroblasts and X-ray-induced aging hepatic stellate cells.82 Our prior investigations have also demonstrated that both free fatty acid-induced senescence in HK-2 cells and replicative senescence result in elevated levels of ANGPTL4 protein expression.35
Within the tumor microenvironment, senescent non-cancer cells exhibit secretory features under stress conditions, contributing to cancer progression and the development of chemotherapy resistance.83 Centrosome amplification is recognized as a pivotal driver in the oncogenesis and advancement of various malignancies. Recent investigations have unveiled that ANGPTL4 is now recognized as a SASP component, functionally linking cellular senescence, ROS-driven oxidative stress, and tissue-invasive behavior. This connection further highlights its involvement in age-related cancer progression and reinforces the regulatory importance of the ROS-HIF axis in ANGPTL4 activity.84 Additionally, scholars have highlighted ANGPTL4 as a critical late-stage regulatory gene product essential for the transition of cells arrested by CDK4/6 inhibitors into senescent cells capable of producing SASP.28 Further clinical and fundamental research is warranted to ascertain whether secreted ANGPTL4 can be delineated as a component of SASP.
In summary, the potential implications of ANGPTL4 in angiogenesis, the ETM process, modulation of oxidative stress, and involvement in SASP offer novel insights and avenues for the exploration and development of therapeutic strategies (Figure 2). ANGPTL4 plays a “double-edged sword” role in aging: on one hand, it may promote pathological remodeling driven by aging; on the other hand, it maintains vascular homeostasis, particularly exerting protective effects in endothelial and cerebral vascular aging.
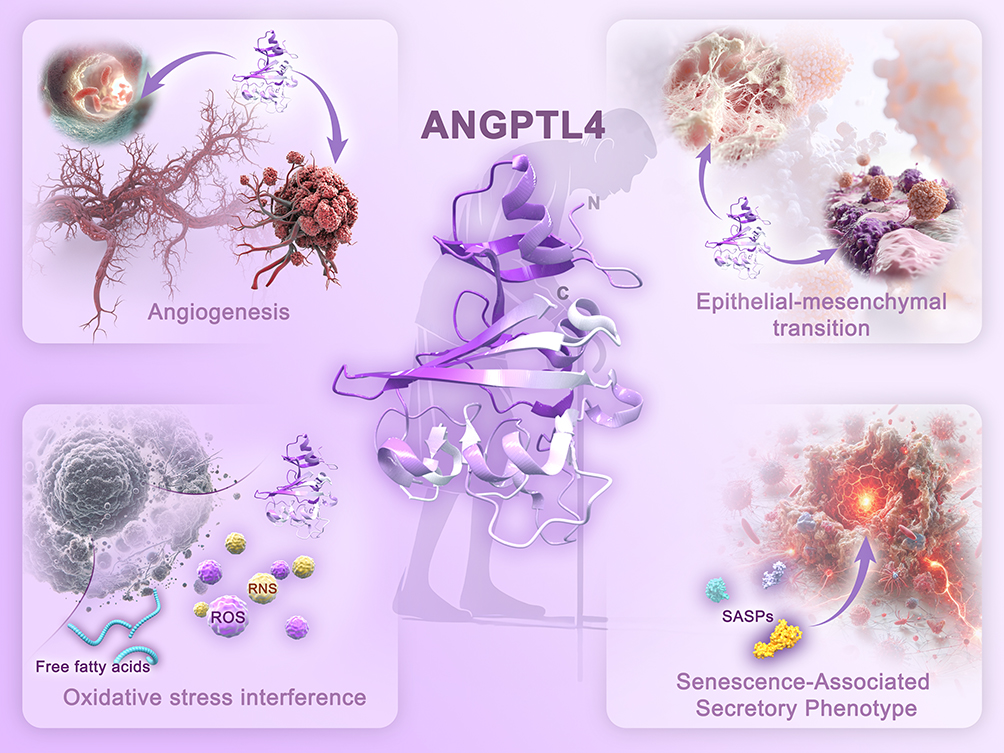 |
Figure 2 The Relationship between ANGPTL4 and Mechanisms Related to Aging. Abbreviations: ROS, Reactive oxygen species; RNS, Reactive nitrogen species; SASP, Senescence-associated secretory phenotype. |
While multiple studies have explored the potential role of ANGPTL4 in the aging process, there are still limitations. Firstly, many studies have not identified the cleavage state of ANGPTL4, and the molecular interaction mechanisms remain unclear. Secondly, tissue specificity and evolutionary conservation need to be verified. Future research can address these issues through a strategy that combines multi-omics integration, computational simulations (molecular docking), and experimental validation (SPR, CRISPR screening).
The Signaling Pathways Involved in ANGPTL4
The oligomerization and protein processing of ANGPTL4 have a significant impact on its in vivo biological activity, and these processes may exert their effects by influencing the interactions of ANGPTL4 with cells and its intracellular signaling pathways.85 Conducting further investigations into the regulatory mechanisms and interactions of the signaling pathways that ANGPTL4 participates in will enhance our comprehension of the underlying mechanisms through which ANGPTL4 operates in the pathogenesis and progression of diseases.
Peroxisome Proliferator-Activated Receptors (PPARs) Signaling Pathway
PPARs participate in regulating processes such as cellular lipid metabolism, inflammatory responses, and cell proliferation, closely associated with aging.86 The ANGPTL4 protein is sensitive to transcriptional control by fatty acids and fatty acid-activated PPARs.11 PPARγ forms a heterodimer with retinoid X receptor (RXR) and can specifically bind to the ANGPTL4 promoter region to enhance its transcriptional activity.87 Activation of PPARα by ligands increases LPL activity by upregulating the expression of GPHBP1 and Lmf1, while reducing the expression of ANGPTL4 protein in proximal tubules, thus mitigating cisplatin-induced nephrotoxicity.88 A study revealed that astrocyte-specific knockout of ANGPTL4 prevented the mitochondrial morphological changes in hypothalamic astrocytes induced by a high-fat diet and increased hypothalamic PPARγ mRNA levels.21 However, in C2C12 myoblasts, the deletion of PPARδ, rather than PPARγ, disrupted the induction of ANGPTL4 mediated by long-chain fatty acids.89 Additionally, research by Adhikary et al has identified the involvement of the PPARβ/δ-ANGPTL4 pathway in the regulation of tumor cell invasion.90
Hypoxia Induced Factor-1α (HIF-1α) Signaling Pathway
HIF-1α is activated in response to hypoxia and plays a crucial role in regulating cellular adaptive responses to low oxygen levels. Dysregulated activation of the HIF-1α signaling pathway can exacerbate cellular aging processes and contribute to overall aging.91 ANGPTL4 is a gene that is induced by hypoxia and is subject to epigenetic regulation.92 Recent studies have unveiled a reciprocal interaction between ANGPTL4 and HIF-1α, suggesting the potential of targeting this pathway as a novel therapeutic approach for renal interstitial fibrosis.93 In a hyperglycemic environment, the upregulation of ANGPTL4 induced by high glucose levels is reliant on the activation of HIF-1α triggered by elevated glucose concentrations both in vivo and in vitro.94 In both in vitro studies involving hypoxic Müller cells and in vivo investigations of the ischemic inner retina, ANGPTL4 is identified as a cytokine that is upregulated by HIF-α. This upregulation plays a critical role in enhancing vascular permeability within hypoxic Müller cells.95 Furthermore, investigations have indicated that the HIF-1α-mediated elevation of ANGPTL4 in adipose tissue, coupled with subsequent inactivation of lipoprotein lipase, may contribute to the development of atherosclerosis in individuals with sleep apnea.96
In the context of tumors, ANGPTL4 expression is prominent in hypoxic regions surrounding necrotic areas.97 Studies by Kang et al propose that nickel-induced accumulation of HIF-1α in lung cells, along with increased levels of ANGPTL4, could be a mechanism through which nickel exposure accelerates the progression of lung cancer.98 Additionally, Li et al have demonstrated that direct upregulation of ANGPTL4 expression by HIF-1α significantly enhances intravascular migration of HCC cells in vitro and promotes liver and distant lung metastases in vivo.99
Extracellular Signal-Regulated Kinase (ERK) Signaling Pathway
The ERK signaling pathway plays a pivotal role in governing cell proliferation, survival, and differentiation, exerting profound effects on cellular functionality and metabolic processes. Hyperactivation of the ERK pathway may disrupt normal cell proliferation and contribute to accelerated aging.100 Fine-tuned regulation of the ERK signaling cascade could potentially ameliorate tissue aging-related pathologies.101 The ERK signaling inhibitor U0126 demonstrates a potent ability to suppress the expression of ANGPTL4.102 Similarly, the inhibition of ERK1/2 signaling transduction with PD98059 significantly attenuates the pro-tumor cell survival function of ANGPTL4.103 Research has elucidated that ANGPTL4 can enhance psoriatic keratinocyte proliferation and inflammatory responses through the ERK1/2-dependent pathway.104 Furthermore, ANGPTL4 instigates the activation of cancer-associated fibroblasts within the tumor microenvironment and facilitates EMT in colorectal cancer cells via the ERK signaling route.105 Moreover, ANGPTL4 plays a crucial role in the proliferation and metastasis of non-small cell lung cancer (NSCLC) cells, partly mediated by the downregulation of the ERK signaling pathway.106 Additionally, the study led by Zhang et al suggests that ANGPTL4 may modulate clear cell renal cell carcinoma cell proliferation through the ERK/P38 pathway.107 In conclusion, a discernible crosstalk exists between ANGPTL4 and the ERK signaling pathway.
Toll-Like Receptor (TLR) Signaling Pathway
The TLR4 signaling pathway is involved in regulating immune responses and inflammatory reactions, and abnormal activation of the TLR4 signaling pathway may promote inflammation and accelerate the aging process.108 Furthermore, the expression of ANGPTL4 is subject to regulation by FFAs, which can stimulate pattern recognition receptors in macrophages, including TLR4.109 In vitro studies have shown that TLR3/4 activation notably enhances the expression and secretion of ANGPTL4 in macrophages.110 Additionally, research has elucidated that the TLR4/MyD88/NF-κB and calcineurin/NFAT signaling cascades are implicated in the upregulation of ANGPTL4 in podocytes following stimulation by lipopolysaccharide (LPS).111 Moreover, LPS-induced activation of the TLR4/IL-22/STAT3 signaling axis influences ANGPTL4 transcription, thereby enhancing lipid absorption in the intestine.112
Moreover, ANGPTL4 likely participates in additional signaling pathways across various diseases. Investigation into these signaling cascades serves to unveil the pivotal roles of ANGPTL4 in cellular biology and disease progression, alongside identifying potential therapeutic targets. Table 1 succinctly outlines the pivotal genes and associated diseases implicated in the aforementioned studies.
 |
Table 1 Key Genes Interacting with ANGPTL4 |
The Potential Application of ANGPTL4 as a Biomarker in Age-Related Diseases
The therapeutic use of antibodies directed against ANGPTL4 has shown promising potential in the management of hyperlipidemia.113 Furthermore, circulating levels of ANGPTL4 have garnered significant clinical research interest in both metabolic disorders and age-related diseases in recent years (Figure 3).
 |
Figure 3 Association of Elevated Circulating ANGPTL4 Levels with Age-Related Diseases. Abbreviations: CKD, chronic kidney disease; CVD, cardiovascular disease; CAP, community-acquired pneumonia; ARDS, acute respiratory distress syndrome. |
Metabolism-Related Diseases
Deactivation of ANGPTL4 can significantly improve glucose tolerance, accompanied by an increase in insulin levels.114 Studies have shown that elevated circulating levels of ANGPTL4 in diabetic patients are associated with inflammation and hypoxia.115 Gene simulations involving ANGPTL4 inhibition and LPL enhancement are associated with lower waist-to-hip ratios, improved insulin glucose metabolism, and a reduced risk of coronary heart disease and type 2 diabetes.116 Furthermore, a clinical study observed that serum ANGPTL4 may act as a crucial link between elevated cholesterol in blood uric acid and triglyceride-rich lipoproteins. The correlation between serum uric acid (SUA) and triglyceride-rich lipoprotein cholesterol (TRL-C) disappeared after controlling for ANGPTL4 levels.117 In another cohort of obese participants, the ANGPTL4-E40K variant can prevent lipid abnormalities.118 The results of the aforementioned clinical studies suggest that the lipid-regulating factor ANGPTL4 holds value for predicting metabolism-related diseases.
Chronic Kidney Disease
Plasma levels of ANGPTL4 show promise as a valuable tool for both identifying and potentially reversing the progression of end-stage renal disease (ESRD).119 Studies have indicated that ANGPTL4 may contribute to kidney injury induced by hyperlipidemia through the downregulation of podocyte ACTN4 expression.120 In the elderly population, serum ANGPTL4 levels exhibit a negative correlation with estimated glomerular filtration rate (eGFR).79 Furthermore, the circulating levels of ANGPTL4 in diabetic patients are significantly correlated with serum creatinine and eGFR levels.121 A cohort study involving 1,115 patients with type 2 diabetes demonstrated that elevated serum ANGPTL4 levels may serve as a clinically significant biomarker for diabetic nephropathy. Notably, mediation analysis revealed that triglycerides do not act as an intermediary factor in the association between ANGPTL4 and nephropathy progression, suggesting a direct or alternative mechanistic pathway linking ANGPTL4 to renal dysfunction in diabetes.122 A retrospective cohort study analysis found that the C allele of rs1044250 and the G allele of rs2278236 in the ANGPTL4 gene are linked to a higher risk of severe proteinuria in kidney transplant patients.123 Utilizing an innovative quantitative flat-panel protein microarray technology, researchers have pinpointed Angptl4, L-selectin, and TGFβ1 in urine as promising candidate biomarkers for monitoring disease activity in lupus nephritis.124
Cardiovascular and Cerebrovascular Diseases
Several observational studies have underscored the potential of ANGPTL4 as a biomarker in cardiovascular disease (CVD).125,126 A recent study has demonstrated a positive correlation between the concentration of C-terminal domain-containing ANGPTL4 (CD-ANGPTL4) and levels of inflammation, the prevalence of diabetes, and cardiovascular mortality.127 Variants of ANGPTL4 have demonstrated possible associations with blood lipid levels and atherosclerotic-related conditions in the Han Chinese population.128 Serum levels of ANGPTL4 have been recognized as an independent risk factor for coronary artery disease (CAD),129 with higher levels of ANGPTL4 linked to a reduced risk of vascular events in acute myocardial infarction patients. The most prevalent functional variant of ANGPTL4, E40K,130 has shown consistent effects on metabolic heart diseases and enhanced lipoprotein lipase activity in recent research.131 Although some studies suggest that the E40K variant may not be independently correlated with plasma ANGPTL4 levels, its significance in cardiovascular diseases remains noteworthy.132
ANGPTL4 has been demonstrated to protect the integrity of the blood-brain barrier in cases of ischemic and thrombolytic injury,133 highlighting its potential as a target for vascular and brain protection in stroke.134 Integrating plasma ANGPTL4 into risk prediction models for acute ischemic stroke patients can enhance the accuracy of predicting combined outcomes such as death and severe disability, indicating the potential utility of ANGPTL4 in assessing stroke risk.135 Additionally, heightened serum levels of ANGPTL4 have been associated with the extent of ischemic stroke lesions, offering insights for further exploration of the role of ANGPTL4 in stroke pathogenesis.136 In a prospective study on chronic obstructive pulmonary disease (COPD) cohorts, elevated levels of ANGPTL4 were independently linked to compromised vascular health and higher NT-proBNP levels.137 In another cohort of rheumatoid arthritis patients, serum ANGPTL4 levels were positively and independently correlated with increased carotid intima-media thickness.138 Moreover, the expression levels of ANGPTL4 are significantly elevated in peripheral blood mononuclear cells (PBMCs) of Alzheimer’s disease (AD) patients, suggesting its potential as a peripheral biomarker for AD.139
Chronic Inflammation-Related Diseases
Chronic inflammation, recognized as a hallmark of the aging process, is intricately linked to aging.140 Clinical studies have indicated that the inflammatory marker CRP can be used to predict plasma levels of ANGPTL4 in individuals, with inflammatory triggers leading to increased expression of ANGPTL4 in macrophages.110 In a separate clinical study, elevated serum levels of ANGPTL4 were observed in patients with acute respiratory distress syndrome (ARDS), showing a close association with inflammatory markers IL-6 and TNF-α.141 The development of a predictive model based on serum ANGPTL4 levels has the potential to assess the severity of community-acquired pneumonia (CAP), paving the way for early interventions in CAP management.142 Research efforts have highlighted the increased expression of ANGPTL4 in the serum and pancreatic tissue of patients with pancreatitis, suggesting that targeting ANGPTL4 could be a promising approach in pancreatitis treatment.143 Moreover, the correlation between elevated serum ANGPTL4 levels and circulating RANKL indicates a potential role for ANGPTL4 as a novel marker for bone degradation in rheumatoid arthritis (RA).144
Tumor Lymph Node Metastasis
ANGPTL4 has been implicated in tumor lymph node metastasis, a critical process in cancer progression.145,146 Tumor cells undergo metabolic reprogramming of lipid metabolism to support metastasis.147 Studies have shown that leptin-induced phosphorylation of ANGPTL4 reduces its interaction with LPL, disrupting lipid homeostasis by elevating intracellular arachidonic acid levels, stimulating prostaglandin E2 (PGE2) production, and facilitating lymph node metastasis in gastric cancer.148 Additionally, ANGPTL4 has been associated with promoting an invasive cancer phenotype by upregulating the expression of matrix metalloproteinases (MMPs).149 Clinical cohort analyses have revealed that serum levels of ANGPTL4 exhibit superior predictive efficacy for vascular invasion and lymph node metastasis in cholangiocarcinoma patients compared to traditional markers such as CA199 and CEA.146 In an in vivo experiment, increased expression of ANGPTL4 in GSAS/N5 cells injected into the tongues of nude mice and subsequently harvested from metastasized lungs demonstrated heightened migratory activity.150
Notably, the different isoforms of ANGPTL4 play contrasting roles in cancer progression. While cANGPTL4 has been shown to promote tumor growth and metastasis, nANGPTL4 exerts inhibitory effects on metastasis by modulating Wnt signaling and reducing vascularization at metastatic sites, thereby enhancing overall survival rates in postoperative metastatic models.53 These findings suggest that ANGPTL4, as a potential novel biomarker, may aid in improving the accuracy of evaluating disease advancement and metastatic risk in cancer patients.
Taken together, as a multifunctional regulatory protein, ANGPTL4 is not only involved in age-related metabolic disorders and chronic inflammatory processes but also closely associated with the progression of various diseases. This dual role endows it with predictive value as a biomarker and intervention potential for targeted therapy. Compared to other aging markers, the core difference of ANGPTL4 lies in its multifunctionality and dynamic secretion characteristics. These features may render it more sensitive in metabolic-related aging diseases than traditional inflammatory markers. However, ANGPTL4 levels may also rise during acute injury or infection, necessitating clinical context interpretation to improve specificity. Our preliminary clinical studies suggest that combined analysis of ANGPTL4 with traditional aging markers may enhance predictive value for frailty status. Future validation through large-scale longitudinal cohort studies is needed to further confirm the independent predictive value of ANGPTL4.
Although targeted therapy for ANGPTL4 shows promise for translation, achieving a balance between efficacy and safety necessitates the use of selective inhibition techniques and precision medicine strategies, including patient stratification and local delivery. Currently, priority should be placed on advancing mechanistic research and validating early clinical trials.
Limitations of Current Clinical Studies
Despite the extensive focus of numerous clinical observational studies on the predictive value of circulating ANGPTL4 in age-related disease occurrence, the current body of evidence lacks substantial support from large-scale cohorts or prospective longitudinal data (Table 2). Moreover, the current studies are predominantly conducted at single centers, primarily concentrating on Asian and European populations, resulting in a notable scarcity of data from African, Latino, and other populations, which could potentially restrict the generalizability of findings. Finally, a majority of studies have not stratified data by gender, overlooking potential gender disparities in the function of ANGPTL4 in lipid metabolism.
 |
Table 2 Clinical Studies of Circulating ANGPTL4 Levels |
Concluding Remarks and Perspectives
At present, the causal relationship between ANGPTL4 dysregulation and the aging phenotype remains unclear. The mechanistic roles of nANGPTL4 and cANGPTL4 in age-related disease pathogenesis and progression are yet to be fully delineated across experimental and clinical studies. This significantly impedes the clinical application of ANGPTL4 antibody therapy. Of particular note is the functional antagonism exhibited by the nANGPTL4 and cANGPTL4 fragments, presenting a significant challenge for intervention strategies based on ANGPTL4. Future therapeutic developments should focus on the development of fragment-specific targeting techniques, such as selectively inhibiting protein cleavage processes or designing conformation-specific antibodies, rather than employing global regulatory strategies.
Circulating levels of ANGPTL4 may have the potential to serve as clinical biomarkers for age-related diseases. However, the current clinical research findings may lack generalizability, and the understanding of the association between ANGPTL4 and aging is still in its early stages. Further prospective clinical and experimental studies are warranted to validate its mechanism of action and potential clinical efficacy. Moreover, the presence of gender disparities151 and tissue-specific variations152–154 in ANGPTL4 levels and their influence on lipid metabolism could offer pivotal insights for the development of individualized therapeutic approaches. While ANGPTL4 shows significant potential in the field of aging-related disease regulation, existing clinical evidence, particularly data pertaining to the elderly population, still has notable limitations. This necessitates a cautious approach when interpreting its biological significance and clinical value.
Abbreviation
FIAF, Fasting-induced adipose factor; ANGPTL4, Angiopoietin-like 4; CCD, coiled-coil domain; FLD, fibrinogen-like domain; LPL, Lipoprotein lipase; PC, Proprotein convertase; AMPK, AMP-activated protein kinase; cANGPTL4, C-terminal fragment ANGPTL4; EMT, Epithelial-mesenchymal transition; SASP, Senescence-associated secretory phenotype; HFpEF, Heart failure with preserved ejection fraction; HCC, Hepatocellular carcinoma; ROS, Reactive oxygen species; RNS, Reactive nitrogen species; FFA, Free fatty acid; PPARs, Peroxisome proliferator-activated receptors; HIF-1α, Hypoxia induced factor-1α; RXR, Retinoid X receptor; ERK, Extracellular signal-regulated kinase; NSCLC, Non-small cell lung cancer; TLR, Toll-like receptor; LPS, Lipopolysaccharide; SUA, Serum uric acid; TRL-C, Triglyceride-rich lipoprotein cholesterol; ESRD, end-stage renal disease; eGFR, estimated glomerular filtration rate; CVD, cardiovascular disease; CD-ANGPTL4, C-terminal domain-containing ANGPTL4; CAD, coronary artery disease; COPD, chronic obstructive pulmonary disease; PBMCs, peripheral blood mononuclear cells; AD, Alzheimer’s disease; ARDS, acute respiratory distress syndrome; CAP, community-acquired pneumonia; RA, rheumatoid arthritis; PGE2, prostaglandin E2; MMPs, matrix metalloproteinases; nANGPTL4, N-terminal fragment of ANGPTL4.
Data Sharing Statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
Funding
This work was supported by the National Natural Science Foundation of China [NO. 82474417] and the Shanghai Medical Leadership Training Program [NO.2019LJ09].
Disclosure
The authors declare that they have no conflicts of interest.
References
1. Lei X, Shi F, Basu D, et al. Proteolytic processing of angiopoietin-like protein 4 by proprotein convertases modulates its inhibitory effects on lipoprotein lipase activity. J Biol Chem. 2011;286(18):15747–15756. doi:10.1074/jbc.M110.217638
2. Biterova E, Esmaeeli M, Alanen H, Saaranen M, Ruddock L. Structures of Angptl3 and Angptl4, modulators of triglyceride levels and coronary artery disease. Sci Rep. 2018;8(1):6752. doi:10.1038/s41598-018-25237-7
3. John J, Richard E, Alexander P, et al. Highly accurate protein structure prediction with AlphaFold. Nature. 2021;596(7873):583.
4. Mihaly V, Stephen A, Mandar D, et al. AlphaFold protein structure database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2021;50:D439–D444.
5. Mandard S, Zandbergen F, van Straten E, et al. The fasting-induced adipose factor/angiopoietin-like protein 4 is physically associated with lipoproteins and governs plasma lipid levels and adiposity. J Biol Chem. 2006;281(2):934–944. doi:10.1074/jbc.M506519200
6. Mandard S, Zandbergen F, Tan N, et al. The direct peroxisome proliferator-activated receptor target fasting-induced adipose factor (FIAF/PGAR/ANGPTL4) is present in blood plasma as a truncated protein that is increased by fenofibrate treatment. J Biol Chem. 2004;279(33):34411–34420. doi:10.1074/jbc.M403058200
7. Allison EM, Deepthi K, Kimberly Y, et al. The C-terminal fibrinogen-like domain of angiopoietin-like 4 stimulates adipose tissue lipolysis and promotes energy expenditure. J Biol Chem. 2017;292(39):16122–16134.
8. Lin J, Naot D, Watson M, et al. Skeletal actions of fasting-induced adipose factor (FIAF). Endocrinology. 2013;154(12):4685–4694. doi:10.1210/en.2013-1238
9. Gunn K, Gutgsell A, Xu Y, Johnson C, Liu J, Neher S. Comparison of angiopoietin-like protein 3 and 4 reveals structural and mechanistic similarities. J Biol Chem. 2021;296:100312. doi:10.1016/j.jbc.2021.100312
10. Baranowski T, Kralisch S, Bachmann A, et al. Serum levels of the adipokine fasting-induced adipose factor/angiopoietin-like protein 4 depend on renal function. Hormone Metab Res. 2011;43(2):117–120. doi:10.1055/s-0030-1267917
11. Wieneke D, Sander K. Regulation of lipoprotein lipase by Angptl4. Trends Endocrinol Metab. 2014;25(3):146–155.
12. Leth-Espensen K, Kristensen K, Kumari A, et al. The intrinsic instability of the hydrolase domain of lipoprotein lipase facilitates its inactivation by ANGPTL4-catalyzed unfolding. Proc Natl Acad Sci USA. 2021;118(12). doi:10.1073/pnas.2026650118.
13. Elena M, Evelina V, Thomas O, Gunilla O. Inactivation of lipoprotein lipase in 3T3-L1 adipocytes by angiopoietin-like protein 4 requires that both proteins have reached the cell surface. Biochem Biophys Res Commun. 2013;441(4):941–946.
14. Xia L, Fujun S, Debapriya B, et al. Proteolytic processing of angiopoietin-like protein 4 by proprotein convertases modulates its inhibitory effects on lipoprotein lipase activity. J Biol Chem. 2011;286(18):9577–9586. doi:10.1074/jbc.M110.208777
15. Kersten S, Lichtenstein L, Steenbergen E, et al. Caloric restriction and exercise increase plasma ANGPTL4 levels in humans via elevated free fatty acids. Arteriosclerosis Thrombosis Vasc Biol. 2009;29(6):969–974. doi:10.1161/ATVBAHA.108.182147
16. Astrid Johannesson H, Esben S, Steen Bønløkke P, Niels M, Niels J, Jens Otto LJ. Growth hormone upregulates ANGPTL4 mRNA and suppresses lipoprotein lipase via fatty acids: randomized experiments in human individuals. Metabolism. 2020;105:154188.
17. Ingerslev B, Hansen J, Hoffmann C, et al. Angiopoietin-like protein 4 is an exercise-induced hepatokine in humans, regulated by glucagon and cAMP. Mol Metabol. 2017;6(10):1286–1295. doi:10.1016/j.molmet.2017.06.018
18. Chang H, Kwon O, Shin M, et al. Role of Angptl4/Fiaf in exercise-induced skeletal muscle AMPK activation. J Appl Physiol. 2018;125(3):715–722. doi:10.1152/japplphysiol.00984.2016
19. Dijk W, Heine M, Vergnes L, et al. ANGPTL4 mediates shuttling of lipid fuel to brown adipose tissue during sustained cold exposure. eLife. 2015;4:e08428.
20. Kim H, Youn B, Shin M, et al. Hypothalamic Angptl4/Fiaf is a novel regulator of food intake and body weight. Diabetes. 2010;59(11):2772–2780. doi:10.2337/db10-0145
21. Luis V, Jae Geun K, Pablo F-T, et al. Astrocytic lipid metabolism determines susceptibility to diet-induced obesity. Sci Adv. 2021;7(50):eabj2814.
22. Flores-León M, Alcaraz N, Pérez-Domínguez M, et al. Transcriptional profiles reveal deregulation of lipid metabolism and inflammatory pathways in neurons exposed to palmitic acid. Mol Neurobiol. 2021;58(9):4639.
23. Vienberg S, Kleinridders A, Suzuki R, Kahn C. Differential effects of angiopoietin-like 4 in brain and muscle on regulation of lipoprotein lipase activity. Mol Metabol. 2015;4(2):144–150. doi:10.1016/j.molmet.2014.11.003
24. Mattijssen F, Alex S, Swarts H, Groen A, van Schothorst E, Kersten S. Angptl4 serves as an endogenous inhibitor of intestinal lipid digestion. Mol Metabol. 2014;3(2):135–144. doi:10.1016/j.molmet.2013.11.004
25. Alex S, Lichtenstein L, Dijk W, Mensink R, Tan N, Kersten S. ANGPTL4 is produced by entero-endocrine cells in the human intestinal tract. Histochem Cell Biol. 2014;141(4):383–391. doi:10.1007/s00418-013-1157-y
26. Janssen A, Dijk W, Boekhorst J, et al. ANGPTL4 promotes bile acid absorption during taurocholic acid supplementation via a mechanism dependent on the gut microbiota. Biochim Biophys Acta Mol Cell Biol Lipids. 2017;1862(10):1056–1067. doi:10.1016/j.bbalip.2017.07.005
27. Hernández-Rocha C, Borowski K, Turpin W, et al. Integrative analysis of colonic biopsies from inflammatory bowel disease patients identifies an interaction between microbial bile acid-inducible gene abundance and human angiopoietin-like 4 gene expression. J Crohn’s Colitis. 2021;15(12):2078–2087. doi:10.1093/ecco-jcc/jjab096
28. Gleason C, Dickson M, Klein Dooley M, et al. Therapy-induced senescence contributes to the efficacy of abemaciclib in patients with dedifferentiated liposarcoma. Clin Cancer Res. 2024;30(4):703–718. doi:10.1158/1078-0432.CCR-23-2378
29. Qi J, Ruolan M, Yuhuan W, et al. ANGPTL4 inhibits granulosa cell proliferation in polycystic ovary syndrome by EGFR/JAK1/STAT3-mediated induction of p21. FASEB J. 2023;37(2). doi:10.1096/fj.202201796RR.
30. L’Episcopo F, Tirolo C, Testa N, et al. Wnt/β-catenin signaling is required to rescue midbrain dopaminergic progenitors and promote neurorepair in ageing mouse model of Parkinson’s disease. Stem Cells. 2014;32(8):2147–2163. doi:10.1002/stem.1708
31. Jeoung J, Nam H, Kwak J, et al. A decline in Wnt3a signaling is necessary for mesenchymal stem cells to proceed to replicative senescence. Stem Cells and Development. 2015;24(8):973–982. doi:10.1089/scd.2014.0273
32. Kirsch N, Chang L, Koch S, et al. Angiopoietin-like 4 is a Wnt signaling antagonist that promotes LRP6 turnover. Dev Cell. 2017;43(1):71–82.e6. doi:10.1016/j.devcel.2017.09.011
33. Son Y, Lorenz W, Paton C. Linoleic acid-induced ANGPTL4 inhibits C2C12 skeletal muscle differentiation by suppressing Wnt/β-catenin. J Nutr Biochem. 2023;116:109324. doi:10.1016/j.jnutbio.2023.109324
34. Palikaras K, Mari M, Ploumi C, Princz A, Filippidis G, Tavernarakis N. Age-dependent nuclear lipid droplet deposition is a cellular hallmark of aging in Caenorhabditis elegans. Aging Cell. 2023;22(4):e13788. doi:10.1111/acel.13788
35. Wang X, Chang H, Gu X, et al. Renal lipid accumulation and aging linked to tubular cells injury via ANGPTL4. Mechanisms Ageing Develop. 2024;219:111932. doi:10.1016/j.mad.2024.111932
36. Wen Y, Zhang X, Liu H, et al. SGLT2 inhibitor downregulates ANGPTL4 to mitigate pathological aging of cardiomyocytes induced by type 2 diabetes. Cardiovas Diabetol. 2024;23(1):430. doi:10.1186/s12933-024-02520-8
37. Ghosh A, Chénier I, Leung Y, et al. Adipocyte Angptl8 deletion improves glucose and energy metabolism and obesity associated inflammation in mice. iScience. 2024;27(12):111292. doi:10.1016/j.isci.2024.111292
38. Kersten S. ANGPTL3 as therapeutic target. Curr Opin Lipidol. 2021;32(6):335–341. doi:10.1097/MOL.0000000000000789
39. Carlos L-O, Maria AB, Linda P, Manuel S, Guido K. Hallmarks of aging: an expanding universe. Cell. 2023. 186(2):243–278.
40. Burcu S, Brian IC, Hani A. EMT and inflammation: crossroads in HCC. J Gastrointest Cancer. 2022;54(1):204–212.
41. Ana Karina A-R, Alfredo C-G, Omar Emiliano A-T, José P-C. Mitochondrial redox signaling and oxidative stress in kidney diseases. Biomolecules. 2021;11(8):1144.
42. Mir Mohd F, Nathan DS, Syed Mudabir A, et al. Convergence of therapy-induced senescence (TIS) and EMT in multistep carcinogenesis: current opinions and emerging perspectives. Cell Death Discov. 2020;6:51.
43. Simone B, Rafael AED, Ling H, Maxim NS, Martin WH. Transcriptional and functional changes of the human microvasculature during physiological aging and Alzheimer disease. Adv Biosyst. 2020;4(5):2000044.
44. Ananya C, Alwin K, Bert V, et al. Angiopoietin like-4 as a novel vascular mediator in capillary cerebral amyloid angiopathy. Brain. 2018;141(12):3377–3388.
45. Perdiguero E, Galaup A, Durand M, et al. Alteration of developmental and pathological retinal angiogenesis in angptl4-deficient mice. J Biol Chem. 2011;286(42):36841–36851. doi:10.1074/jbc.M111.220061
46. Zhandong Q, Jia Y, Gang D, Dayong L, Suming Z. Angiopoietin-like 4 promotes angiogenesis and neurogenesis in a mouse model of acute ischemic stroke. Brain Res Bull. 2021;168:156–164.
47. Mousavizadeh R, Scott A, Lu A, et al. Angiopoietin-like 4 promotes angiogenesis in the tendon and is increased in cyclically loaded tendon fibroblasts. J Physiol. 2016;594(11):2971–2983. doi:10.1113/JP271752
48. Gomez Perdiguero E, Liabotis-Fontugne A, Durand M, et al. ANGPTL4-αvβ3 interaction counteracts hypoxia-induced vascular permeability by modulating Src signalling downstream of vascular endothelial growth factor receptor 2. J Pathol. 2016;240(4):461–471. doi:10.1002/path.4805
49. Chaube B, Citrin K, Sahraei M, et al. Suppression of angiopoietin-like 4 reprograms endothelial cell metabolism and inhibits angiogenesis. Nat Commun. 2023;14(1):8251. doi:10.1038/s41467-023-43900-0
50. Li Y, Gao A, Zeng T, et al. ANGPTL4 accelerates ovarian serous cystadenocarcinoma carcinogenesis and angiogenesis in the tumor microenvironment by activating the JAK2/STAT3 pathway and interacting with ESM1. J Transl Med. 2024;22(1):46. doi:10.1186/s12967-023-04819-8
51. Kolb R, Kluz P, Tan Z, et al. Obesity-associated inflammation promotes angiogenesis and breast cancer via angiopoietin-like 4. Oncogene. 2019;38(13):2351–2363. doi:10.1038/s41388-018-0592-6
52. Ying-Hua Y, Yu W, Karen SLL, et al. Suppression of the Raf/MEK/ERK signaling cascade and inhibition of angiogenesis by the carboxyl terminus of angiopoietin-like protein 4. Arterioscler Thromb Vasc Biol. 2008;28(5):835.
53. Corinne H, Ashik Ahmed AP, Denise G, et al. Primary tumor-derived systemic nANGPTL4 inhibits metastasis. J Exp Med. 2022;220(1):e20202595.
54. Royston-Luke H, Ziqiang T, Han Chung C, et al. ANGPTL4 modulates vascular junction integrity by integrin signaling and disruption of intercellular VE-cadherin and claudin-5 clusters. Blood. 2011;118(14):3990–4002.
55. Jiali Y, Juan L, Jiayu L, et al. Epithelial-mesenchymal transition in age-associated thymic involution: mechanisms and therapeutic implications. Ageing Res Rev. 2023;92:102115.
56. Xue-feng X, Hua-ping D. Type 2 epithelial mesenchymal transition in vivo: truth or pitfalls? Chin Med J. 2012;125(18):3312–3317.
57. Saito S, Kitabatake M, Ouji-Sageshima N, et al. Angiopoietin-like 4 is a critical regulator of fibroblasts during pulmonary fibrosis development. Am J Respir Cell Mol Biol. 2023;69(3):328–339. doi:10.1165/rcmb.2022-0304OC
58. Li G, Zhao H, Cheng Z, Liu J, Li G, Guo Y. Single-cell transcriptomic profiling of heart reveals ANGPTL4 linking fibroblasts and angiogenesis in heart failure with preserved ejection fraction. J Adv Res. 2024;68:215–230.
59. Xin Z, Shanshan Y, Xin Z, et al. ANGPTL4 regulates CD163 expression and Kuppfer cell polarization induced cirrhosis via TLR4/NF-κB pathway. Exp Cell Res. 2021;405(2):112706.
60. Valiakou V, Eliadis P, Karamichali E, et al. Differential expression of the host lipid regulators ANGPTL-3 and ANGPTL-4 in HCV infection and treatment. Int J Mol Sci. 2021;22(15):7961. doi:10.3390/ijms22157961
61. Ke L, He Q, Qu J, et al. Bone-protective effects of neutralizing angiopoietin-like protein 4 monoclonal antibody in rheumatoid arthritis. Mol Ther. 2024;32(12):4497–4513. doi:10.1016/j.ymthe.2024.09.031
62. Ievgenia P, Cédric B. EMT transition states during tumor progression and metastasis. Trends Cell Biol. 2018;29(3):212–226.
63. Brabletz S, Schuhwerk H, Brabletz T, Stemmler M. Dynamic EMT: a multi-tool for tumor progression. EMBO J. 2021;40(18):e108647. doi:10.15252/embj.2021108647
64. Liao Z, Lim J, Lee J, et al. Attenuating epithelial-to-mesenchymal transition in cancer through angiopoietin-like 4 inhibition in a 3D tumor microenvironment model. Adv Healthcare Mater. 2024;13(10):e2303481. doi:10.1002/adhm.202303481
65. Gordon E, Wright C, James M, Cooper S. Transcriptomic and functional analysis of ANGPTL4 overexpression in pancreatic cancer nominates targets that reverse chemoresistance. BMC Cancer. 2023;23(1):524. doi:10.1186/s12885-023-11010-1
66. Shen C, Chan S, Lee C, Huang W, Tsai J, Chen B. Oleic acid-induced ANGPTL4 enhances head and neck squamous cell carcinoma anoikis resistance and metastasis via up-regulation of fibronectin. Cancer Lett. 2017;386:110–122. doi:10.1016/j.canlet.2016.11.012
67. Teo Z, Sng M, Chan J, et al. Elevation of adenylate energy charge by angiopoietin-like 4 enhances epithelial-mesenchymal transition by inducing 14-3-3γ expression. Oncogene. 2017;36(46):6408–6419. doi:10.1038/onc.2017.244
68. Stephen FV, Jie Z, Marko O, Tolga B, Rotem N, Dorothy EV. Healthful aging mediated by inhibition of oxidative stress. Ageing Res Rev. 2020;64:101194.
69. Filomeni G, De Zio D, Cecconi F. Oxidative stress and autophagy: the clash between damage and metabolic needs. Cell Death Differ. 2014;22(3):377–388.
70. Catoire M, Alex S, Paraskevopulos N, et al. Fatty acid-inducible ANGPTL4 governs lipid metabolic response to exercise. Proc Natl Acad Sci USA. 2014;111(11):E1043–52. doi:10.1073/pnas.1400889111
71. Grandl G, Wolfrum C. Hemostasis, endothelial stress, inflammation, and the metabolic syndrome. Semin Immunopathol. 2018;40(2):215–224. doi:10.1007/s00281-017-0666-5
72. Abhishek KS, Balkrishna C, Xinbo Z, et al. Hepatocyte-specific suppression of ANGPTL4 improves obesity-associated diabetes and mitigates atherosclerosis in mice. J Clin Invest. 2021;131(17).
73. Anastasia G, Laeticia L, Tatjana D, et al. Induction of cardiac Angptl4 by dietary fatty acids is mediated by peroxisome proliferator-activated receptor beta/delta and protects against fatty acid-induced oxidative stress. Circ Res. 2010;106(11):1712–1721.
74. Legrand-Poels S, Esser N, L’homme L, Scheen A, Paquot N, Piette J. Free fatty acids as modulators of the NLRP3 inflammasome in obesity/type 2 diabetes. Biochem Pharmacol. 2014;92(1):131–141. doi:10.1016/j.bcp.2014.08.013
75. Qingbin C, Jing-Quan W, Yehuda GA, et al. Modulating ROS to overcome multidrug resistance in cancer. Drug Resist Updat. 2018;41:1–25.
76. Pengcheng Z, Ming Jie T, Royston-Luke H, et al. Angiopoietin-like 4 protein elevates the prosurvival intracellular O2(-):H2O2 ratio and confers anoikis resistance to tumors. Cancer Cell. 2011;19(3):401–415.
77. Chih-Jie S, Kwang-Yu C, Bo-Wen L, et al. Oleic acid-induced NOX4 is dependent on ANGPTL4 expression to promote human colorectal cancer metastasis. Theranostics. 2020;10(16):7083.
78. Tchkonia T, Zhu Y, van Deursen J, Campisi J, Kirkland J. Cellular senescence and the senescent secretory phenotype: therapeutic opportunities. J Clin Invest. 2013;123(3):966–972. doi:10.1172/JCI64098
79. Hung-Chen C, Xiaojun W, Xuchao G, et al. Correlation of serum VEGF-C, ANGPTL4, and activin A levels with frailty. Exp Gerontol. 2023;185:112345.
80. Park S, Min E, Kim S, et al. Exploring memory function beyond immune cells: ANGPTL4-mediated memory functions in tissue resident stem cells. Adv Sci. 2024;11(28):e2307545. doi:10.1002/advs.202307545
81. Nancy A, Courtney H, Barbara Dotto F, Samuel CR, William HJN, Matthew OP. Angptl4 gene expression as a marker of adaptive homeostatic response to social isolation across the lifespan in zebrafish. Neurobiol Aging. 2023;131:209–221.
82. Odagiri N, Matsubara T, Higuchi M, et al. Involvement of ERK1/2 activation in the gene expression of senescence-associated secretory factors in human hepatic stellate cells. Mol Cell Biochem. 2019;455(1–2):7–19. doi:10.1007/s11010-018-3466-x
83. Tadahito Y, Mayu K, Atsuko Y, et al. Inflammation-driven senescence-associated secretory phenotype in cancer-associated fibroblasts enhances peritoneal dissemination. Cell Rep. 2021;34(8).
84. Selwin KW, Juliana A, Shu Chian T, Remigio P. The variant senescence-associated secretory phenotype induced by centrosome amplification constitutes a pathway that activates hypoxia-inducible factor-1α. Aging Cell. 2023;22(3):e13766.
85. Ge H, Yang G, Huang L, Motola D, Pourbahrami T, Li C. Oligomerization and regulated proteolytic processing of angiopoietin-like protein 4. J Biol Chem. 2004;279(3):2038–2045. doi:10.1074/jbc.M307583200
86. Erol A. The functions of PPARs in aging and longevity. PPAR Res. 2007;2007:39654. doi:10.1155/2007/39654
87. Lei L, Xu Z, Meng J, Fei G, Qin F, Jianhua L. ANGPTL4 mediates the protective role of PPARγ activators in the pathogenesis of preeclampsia. Cell Death Dis. 2017;8(9):e3054.
88. Li S, Nagothu K, Ranganathan G, et al. Reduced kidney lipoprotein lipase and renal tubule triglyceride accumulation in cisplatin-mediated acute kidney injury. Am J Physiol Renal Physiol. 2012;303(3):F437–48. doi:10.1152/ajprenal.00111.2012
89. Staiger H, Haas C, Machann J, et al. Muscle-derived angiopoietin-like protein 4 is induced by fatty acids via peroxisome proliferator-activated receptor (PPAR)-delta and is of metabolic relevance in humans. Diabetes. 2009;58(3):579–589. doi:10.2337/db07-1438
90. Adhikary T, Brandt D, Kaddatz K, et al. Inverse PPARβ/δ agonists suppress oncogenic signaling to the ANGPTL4 gene and inhibit cancer cell invasion. Oncogene. 2013;32(44):5241–5252. doi:10.1038/onc.2012.549
91. Gao H, Nepovimova E, Heger Z, et al. Role of hypoxia in cellular senescence. Pharmacol Res. 2023;194:106841. doi:10.1016/j.phrs.2023.106841
92. Kang Y, Li C, Tang S, et al. Nickel chloride regulates ANGPTL4 via the HIF-1α-mediated TET1 expression in lung cells. Toxicol Lett. 2021;352:17–25. doi:10.1016/j.toxlet.2021.09.007
93. Li Y, Chen S, Yang Q, et al. The ANGPTL4-HIF-1α loop: a critical regulator of renal interstitial fibrosis. J Transl Med. 2024;22(1):649. doi:10.1186/s12967-024-05466-3
94. Qianyi L, Peirong L, Wei C, Li L, Zhi Z. ANGPTL-4 induces diabetic retinal inflammation by activating Profilin-1. Exp Eye Res. 2017;166:140–150.
95. Xiaoban X, Murilo R, Mahaa U, et al. Hypoxic retinal Muller cells promote vascular permeability by HIF-1-dependent up-regulation of angiopoietin-like 4. Proc Natl Acad Sci U S A. 2013;110(36):E3425–E3434.
96. Luciano FD, Qiaoling Y, Karen LH, et al. Chronic intermittent hypoxia induces atherosclerosis via activation of adipose angiopoietin-like 4. Am J Respir Crit Care Med. 2013;188(2):240–248.
97. Le Jan S, Amy C, Cazes A, et al. Angiopoietin-like 4 is a proangiogenic factor produced during ischemia and in conventional renal cell carcinoma. Am J Pathol. 2003;162(5):1521–1528. doi:10.1016/S0002-9440(10)64285-X
98. Kang Y, Hsu W, Ou C, et al. Metformin mitigates nickel-elicited angiopoietin-like protein 4 expression via HIF-1α for lung tumorigenesis. Int J Mol Sci. 2020;21(2):619.
99. Li H, Ge C, Zhao F, et al. Hypoxia-inducible factor 1 alpha-activated angiopoietin-like protein 4 contributes to tumor metastasis via vascular cell adhesion molecule-1/integrin β1 signaling in human hepatocellular carcinoma. Hepatology. 2011;54(3):910–919. doi:10.1002/hep.24479
100. Cagnol S, Chambard J. ERK and cell death: mechanisms of ERK-induced cell death--apoptosis, autophagy and senescence. FEBS J. 2010;277(1):2–21. doi:10.1111/j.1742-4658.2009.07366.x
101. Widjaja A, Lim W, Viswanathan S, et al. Inhibition of IL-11 signalling extends mammalian healthspan and lifespan. Nature. 2024;632(8023):157–165. doi:10.1038/s41586-024-07701-9
102. Katanasaka Y, Kodera Y, Kitamura Y, Morimoto T, Tamura T, Koizumi F. Epidermal growth factor receptor variant type III markedly accelerates angiogenesis and tumor growth via inducing c-myc mediated angiopoietin-like 4 expression in malignant glioma. Mol Cancer. 2013;12(1):31. doi:10.1186/1476-4598-12-31
103. Xu J, Wu F, Zhu Y, et al. ANGPTL4 regulates ovarian cancer progression by activating the ERK1/2 pathway. Can Cell Inter. 2024;24(1):54. doi:10.1186/s12935-024-03246-z
104. Zuo Y, Dai L, Li L, et al. viaANGPTL4 regulates psoriasis modulating hyperproliferation and inflammation of keratinocytes. Front Pharmacol. 2022;13:850967. doi:10.3389/fphar.2022.850967
105. Kunning Z, Zhiwei Z, Sanshui Y, Yu T. DNA methylation mediated down-regulation of ANGPTL4 promotes colorectal cancer metastasis by activating the ERK pathway. J Cancer. 2021;12(18):5473.
106. Zhu X, Guo X, Wu S, Wei L. ANGPTL4 correlates with NSCLC progression and regulates epithelial-mesenchymal transition via ERK pathway. Lung. 2016;194(4):637–646. doi:10.1007/s00408-016-9895-y
107. Ma B, Qin L, Sun Z, et al. The single-cell evolution trajectory presented different hypoxia heterogeneity to reveal the carcinogenesis of genes in clear cell renal cell carcinoma: based on multiple omics and real experimental verification. Environ Toxicol. 2024;39(2):869–881. doi:10.1002/tox.24009
108. Kim H, Kim H, Lee J, Hwangbo C. Toll-like receptor 4 (TLR4): new insight immune and aging. Immunity Ageing. 2023;20(1):67. doi:10.1186/s12979-023-00383-3
109. Frommer K, Schäffler A, Rehart S, Lehr A, Müller-Ladner U, Neumann E. Free fatty acids: potential proinflammatory mediators in rheumatic diseases. Ann Rheumatic Dis. 2015;74(1):303–310. doi:10.1136/annrheumdis-2013-203755
110. Tjeerdema N, Georgiadi A, Jonker J, et al. Inflammation increases plasma angiopoietin-like protein 4 in patients with the metabolic syndrome and type 2 diabetes. BMJ Open Diabetes Res Care. 2014;2(1):e000034. doi:10.1136/bmjdrc-2014-000034
111. Shen X, Weng C, Wang Y, et al. Lipopolysaccharide-induced podocyte injury is regulated by calcineurin/NFAT and TLR4/MyD88/NF-κB signaling pathways through angiopoietin-like protein 4. Genes Dis. 2022;9(2):443–455. doi:10.1016/j.gendis.2020.07.005
112. Rong B, Wu Q, Reiter R, Sun C. The mechanism of oral melatonin ameliorates intestinal and adipose lipid dysmetabolism through reducing Escherichia coli-derived lipopolysaccharide. CMGH. 2021;12(5):1643–1667. doi:10.1016/j.jcmgh.2021.06.024
113. E-Chiang L, Urvi D, Gennady G, et al. Identification of a new functional domain in angiopoietin-like 3 (ANGPTL3) and angiopoietin-like 4 (ANGPTL4) involved in binding and inhibition of lipoprotein lipase (LPL). J Biol Chem. 2009;284(20):13735–13745.
114. Janssen A, Katiraei S, Bartosinska B, Eberhard D, Willems van Dijk K, Kersten S. Loss of angiopoietin-like 4 (ANGPTL4) in mice with diet-induced obesity uncouples visceral obesity from glucose intolerance partly via the gut microbiota. Diabetologia. 2018;61(6):1447–1458. doi:10.1007/s00125-018-4583-5
115. McCulloch L, Bramwell L, Knight B, Kos K. Circulating and tissue specific transcription of angiopoietin-like protein 4 in human Type 2 diabetes. Metabolism. 2020;106:154192. doi:10.1016/j.metabol.2020.154192
116. Filippo C. New therapeutic targets in lipid lowering: high-sugar feeding in infants, lipoprotein(a), and lipoprotein lipase. Eur Heart J. 2021;42(12):1119–1122.
117. Peng Y, Hu D, Luo Q, Peng D. Angiopoietin-like protein 4 may be an interplay between serum uric acid and triglyceride-rich lipoprotein cholesterol. Front Cardiovas Med. 2022;9:863687. doi:10.3389/fcvm.2022.863687
118. Bailetti D, Bertoccini L, Mancina R, et al. ANGPTL4 gene E40K variation protects against obesity-associated dyslipidemia in participants with obesity. Obesity Science Practice. 2019;5(1):83–90. doi:10.1002/osp4.311
119. Jia S, Peng X, Liang L, et al. The study of Angptl4-modulated podocyte injury in IgA nephropathy. Front Physiol. 2020;11:575722. doi:10.3389/fphys.2020.575722
120. Yue L, Wangqiu G, Jing L, et al. Angiopoietin-like protein 4 promotes hyperlipidemia-induced renal injury by down-regulating the expression of ACTN4. Biochem Biophys Res Commun. 2022;595:69–75.
121. Eman AS, Mohamed A-F, Sriraman D, et al. ANGPTL4: a predictive marker for diabetic nephropathy. J Diabetes Res. 2019;2019:4943191.
122. Wang Y, Li K, Yuan S, et al. Angiopoietin-like 4 is a potential biomarker for diabetic kidney disease in type 2 diabetes patients. J Diabetes Invest. 2024;15(12):1763–1772. doi:10.1111/jdi.14304
123. Youngil C, Tariq S, Jaewook Y, David IM. Association of genetic polymorphisms of angiopoietin-like 4 with severity of posttransplant proteinuria in kidney allograft recipients. Transpl Immunol. 2016;40:35–41.
124. Kamala V, Sanam S, Ting Z, et al. Quantitative planar array screen of 1000 proteins uncovers novel urinary protein biomarkers of lupus nephritis. Ann Rheum Dis. 2020;79:1349–1361.
125. Abu-Farha M, Cherian P, Qaddoumi M, et al. Increased plasma and adipose tissue levels of ANGPTL8/Betatrophin and ANGPTL4 in people with hypertension. Lipids Health Dis. 2018;17(1):35. doi:10.1186/s12944-018-0681-0
126. Nowowiejska J, Baran A, Hermanowicz J, et al. Angiopoietin-Like 4 (ANGPTL4) in patients with psoriasis, Lichen Planus and Vitiligo-A Pilot Study from the Bialystok+ Polish Longitudinal University Study. Metabolites. 2022;12(9):877. doi:10.3390/metabo12090877
127. Silbernagel G, Chen Y, Li H, et al. Associations of circulating ANGPTL3, C-terminal domain-containing ANGPTL4, and ANGPTL3/8 and ANGPTL4/8 complexes with LPL activity, diabetes, inflammation, and cardiovascular mortality. Circulation. 2024;151(3):218–234. doi:10.1161/CIRCULATIONAHA.124.069272
128. Yang Q, Yin R, Cao X, Huang F, Zhou Y, Chen W. ANGPTL4 variants and their haplotypes are associated with serum lipid levels, the risk of coronary artery disease and ischemic stroke and atorvastatin cholesterol-lowering responses. Nutr Metab. 2018;15(1):70. doi:10.1186/s12986-018-0308-5
129. Zhao Z, Fu Y, Lian H, et al. Correlation between the serum FABP4, ANGPTL3, and ANGPTL4 levels and coronary artery disease. Clin Cardiol. 2024;47(3):e24246. doi:10.1002/clc.24246
130. Yin W, Romeo S, Chang S, Grishin N, Hobbs H, Cohen J. Genetic variation in ANGPTL4 provides insights into protein processing and function. J Biol Chem. 2009;284(19):13213–13222. doi:10.1074/jbc.M900553200
131. Gagnon E, Bourgault J, Gobeil É, Thériault S, Arsenault B. Impact of loss-of-function in angiopoietin-like 4 on the human phenome. Atherosclerosis. 2024;393:117558. doi:10.1016/j.atherosclerosis.2024.117558
132. Smart-Halajko M, Robciuc M, Cooper J, et al. The relationship between plasma angiopoietin-like protein 4 levels, angiopoietin-like protein 4 genotype, and coronary heart disease risk. Arteriosclerosis Thrombosis Vasc Biol. 2010;30(11):2277–2282. doi:10.1161/ATVBAHA.110.212209
133. Zhang B, Xu X, Chu X, Yu X, Zhao Y. Protective effects of angiopoietin-like 4 on the blood-brain barrier in acute ischemic stroke treated with thrombolysis in mice. Neurosci lett. 2017;645:113–120. doi:10.1016/j.neulet.2017.03.001
134. Bouleti C, Mathivet T, Coqueran B, et al. Protective effects of angiopoietin-like 4 on cerebrovascular and functional damages in ischaemic stroke. Eur Heart J. 2013;34(47):3657–3668. doi:10.1093/eurheartj/eht153
135. Xiaowei Z, Suwen S, Aili W, et al. Angiopoietin-like protein 4 and clinical outcomes in ischemic stroke patients. Ann Clin Transl Neurol. 2021;8(3):687–695.
136. He X, Shen Y, Zhu M, et al. Angiopoietin-like protein 4 serum levels and gene polymorphisms are associated with large artery atherosclerotic stroke. J Neurol Sci. 2016;362:333–338. doi:10.1016/j.jns.2016.02.009
137. Waschki B, Kirsten A, Holz O, et al. Angiopoietin-like protein 4 and cardiovascular function in COPD. BMJ Open Respiratory Res. 2016;3(1):e000161. doi:10.1136/bmjresp-2016-000161
138. de Armas-Rillo L, Quevedo-Abeledo J, Hernández-Hernández V, et al. The angiopoietin-like protein 4, apolipoprotein C3, and lipoprotein lipase axis is disrupted in patients with rheumatoid arthritis. Arthritis Res Therapy. 2022;24(1):99. doi:10.1186/s13075-022-02784-z
139. Leandro G, Evangelista A, Lobo R, Xavier D, Moriguti J, Sakamoto-Hojo E. Changes in expression profiles revealed by transcriptomic analysis in peripheral blood mononuclear cells of Alzheimer’s disease patients. J Alzheimers Dis. 2018;66(4):1483–1495. doi:10.3233/JAD-170205
140. Li X, Li C, Zhang W, Wang Y, Qian P, Huang H. Inflammation and aging: signaling pathways and intervention therapies. Signal Transduct Target Ther. 2023;8(1):239. doi:10.1038/s41392-023-01502-8
141. Jun H, Lian L, Xianghu Z, et al. Prognostic value of angiopoietin-like 4 in patients with acute respiratory distress syndrome. Shock. 2021;56:403–411.
142. Chen S, Jiang J, Su M, et al. A nomogram based on the expression level of angiopoietin-like 4 to predict the severity of community-acquired pneumonia. BMC Infect Dis. 2023;23(1):677. doi:10.1186/s12879-023-08648-4
143. Jung K, Son M, Yan H, et al. ANGPTL4 exacerbates pancreatitis by augmenting acinar cell injury through upregulation of C5a. EMBO Mol Med. 2020;12(8):e11222. doi:10.15252/emmm.201911222
144. Swales C, Athanasou N, Knowles H. Angiopoietin-like 4 is over-expressed in rheumatoid arthritis patients: association with pathological bone resorption. PLoS One. 2014;9(10):e109524. doi:10.1371/journal.pone.0109524
145. Akishima-Fukasawa Y, Ishikawa Y, Akasaka Y, et al. Histopathological predictors of regional lymph node metastasis at the invasive front in early colorectal cancer. Histopathology. 2011;59(3):470–481. doi:10.1111/j.1365-2559.2011.03964.x
146. Tin May A, Mang Ngaih C, Atit S, et al. Serum angiopoietin-like protein 4: a potential prognostic biomarker for prediction of vascular invasion and lymph node metastasis in cholangiocarcinoma patients. Front Public Health. 2022;10:836985.
147. Lee C, Jeong S, Jang C, et al. Tumor metastasis to lymph nodes requires YAP-dependent metabolic adaptation. Science. 2019;363(6427):644–649. doi:10.1126/science.aav0173
148. Jian X, Shuqing C, Jiawei W, et al. Leptin-mediated suppression of lipoprotein lipase cleavage enhances lipid uptake and facilitates lymph node metastasis in gastric cancer. Cancer Commun. 2024;44(8):855–878.
149. William Y, Elie K, Qianyu G, et al. MNK1 signaling induces an ANGPTL4-mediated gene signature to drive melanoma progression. Oncogene. 2020;39(18):3650–3665.
150. Tanaka T, Imamura T, Yoneda M, et al. Enhancement of active MMP release and invasive activity of lymph node metastatic tongue cancer cells by elevated signaling via the TNF-α-TNFR1-NF-κB pathway and a possible involvement of angiopoietin-like 4 in lung metastasis. Int J Oncol. 2016;49(4):1377–1384. doi:10.3892/ijo.2016.3653
151. Deng M, Kersten S. Characterization of sexual dimorphism in ANGPTL4 levels and function. J Lipid Res. 2024;65(4):100526. doi:10.1016/j.jlr.2024.100526
152. Aryal B, Singh A, Zhang X, et al. Absence of ANGPTL4 in adipose tissue improves glucose tolerance and attenuates atherogenesis. JCI Insight. 2018;3(6). doi:10.1172/jci.insight.97918.
153. Singh A, Aryal B, Chaube B, et al. Brown adipose tissue derived ANGPTL4 controls glucose and lipid metabolism and regulates thermogenesis. Mol Metabol. 2018;11:59–69. doi:10.1016/j.molmet.2018.03.011
154. Hu H, Luo S, Lai P, et al. ANGPTL4 binds to the leptin receptor to regulate ectopic bone formation. Proc Natl Acad Sci USA. 2024;121(1):e2310685120. doi:10.1073/pnas.2310685120
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.