")
Back to Journals » Journal of Inflammation Research » Volume 17
Candida albicans-Derived β-Glucan as a Novel Modulator of Tumor Microenvironment: Targeting Macrophage Polarization and Inducing Ferroptosis in Lung Cancer
Authors He X, Ran Q, Li X, Xiong A, Zhang L, Jiang M, Bai L, Peng D, Wang J, Sun B, Li G
Received 29 July 2024
Accepted for publication 23 November 2024
Published 6 December 2024 Volume 2024:17 Pages 10479—10494
DOI https://doi.org/10.2147/JIR.S489191
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Xiang He,1– 3,* Qin Ran,2,3,* Xiaolan Li,2,3,* Anying Xiong,2,3 Lei Zhang,2,3 Manling Jiang,2,3 Lingling Bai,2 Dan Peng,2 Junyi Wang,2,3 Baoqing Sun,1 Guoping Li1– 3
1National Center for Respiratory Medicine, National Clinical Research Center for Respiratory Disease, State Key Laboratory of Respiratory Disease, Guangzhou Institute of Respiratory Health, The First Affiliated Hospital of Guangzhou Medical University, Guangzhou, Guangdong, 510120, People’s Republic of China; 2Laboratory of Allergy and Precision Medicine, Chengdu Institute of Respiratory Health, Affiliated Hospital of Southwest Jiaotong University, The Third People’s Hospital of Chengdu, Chengdu, 610031, People’s Republic of China; 3Department of Respiration, Chengdu third People’s hospital Branch of National Clinical Research Center for Respiratory Disease, Affiliated Hospital of ChongQing Medical University, Chengdu, 610031, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Guoping Li; Baoqing Sun, Email [email protected]; [email protected]
Introduction: Tumor-associated macrophages (TAMs) play a crucial role in the tumor microenvironment (TME), and their polarization state significantly influences patient outcomes. This study investigates the inhibitory effects of β-glucan extracted from Candida albicans on lung cancer progression, focusing on its impact on TAM polarization and the induction of ferroptosis, a form of regulated cell death.
Methods: Utilizing both in vivo animal models and in vitro cellular assays, we assessed the impact of β-glucan on tumor growth, cellular proliferation, and migration. We evaluated TAM polarization by analyzing the expression of M1 and M2 markers and identified differentially expressed genes (DEGs) related to ferroptosis. The role of ferroptosis in TAM polarization was further confirmed by assessing the protein levels of ACSL4 and GPX4, intracellular ferrous ion levels, and lipid peroxides.
Results: β-glucan treatment significantly reduced tumor size and weight, along with cellular proliferation and migration, suggesting a potent suppressive effect on lung cancer cell growth. β-glucan promoted an M1-like phenotype in TAMs, as evidenced by increased CD86 expression and decreased CD206 expression, and modulated cytokine mRNA levels. RNA sequencing analysis post β-glucan treatment identified a substantial number of DEGs enriched in the ferroptosis pathway. The induction of ferroptosis by β-glucan was further confirmed through the significant upregulation of ACSL4 and downregulation of GPX4, alongside increased intracellular ferrous ion levels and lipid peroxides. The ferroptosis inhibitor Fer-1 abrogated these effects, highlighting the specificity of β-glucan-mediated polarization.
Conclusion: These results collectively provide novel insights into the immunotherapeutic potential of β-glucan from Candida albicans and its role in modulating TAM polarization and lung cancer growth, offering a promising avenue for cancer treatment strategies.
Keywords: β-glucan, lung cancer, TAMs, macrophage polarization, ferroptosis, C. albicans
Introduction
Lung cancer is the leading cause of cancer-related mortality worldwide, with approximately 2 million new cases and over 1.76 million deaths annually.1 The therapeutic landscape for lung cancer encompasses a range of strategies including surgery, radiotherapy, chemotherapy, molecular targeted therapies, and increasingly, immunotherapy.2 Among these, immunotherapy has become the first-line treatment for advanced lung cancer.3 Consequently, there is a burgeoning interest in the development of immunotherapeutic agents and adjuncts that can enhance the efficacy of immunotherapy for the treatment.
Tumor-associated macrophages (TAMs) play a crucial role in tumor microenvironment (TME), exerting significant influence over immune responses and being intimately connected to patient outcomes.4–7 In the TME, TAMs often adopt the M2-like state, directly promoting cancer cell proliferation, survival, invasion, and metastasis, and aiding in immune evasion.8 Thus, strategies targeting TAMs, such as genetic and epigenetic reprogramming to foster an anti-tumor phenotype, are at the forefront of lung cancer immunotherapy.9 Ferroptosis, a recently identified form of regulated cell death marked by lipid peroxidation and iron accumulation, is a critical process in immunotherapy, typically modulated by TAMs.10,11 Evidence suggests that macrophages can initiate ferroptosis through various signaling pathways, and the byproducts of this cell death can influence TAM polarization.12–14 Hence, investigating the relationship between ferroptosis and TAMs may provide new insights for the immunotherapy of lung cancer.
β-glucan is a key component of fungal cell walls and uniquely constitutes the entire cell wall of C. albicans.15,16 This biologically active polysaccharide has demonstrated anti-metastatic capabilities and the power to induce trained immunity, which can suppress tumor metastasis.17,18 Importantly, β-glucan does not exhibit direct cytotoxicity to cancer cells or tumors.19 Instead, its effects are mediated through the activation of immune cells, including macrophages and dendritic cells, which secrete pro-inflammatory cytokines. Given this context, there is a pressing need to explore the role of β-glucan from C. albicans in tumor biology. The current study explored the anti-cancer effects of β-glucan extracted from C. albicans, focusing on its potential to modulate TAM polarization and its underlying mechanisms in lung cancer. The detailed exploration of β-glucan’s role in modulating the tumor microenvironment offered insights into a potential new avenue for lung cancer treatment and highlighted the therapeutic potential of targeting macrophage polarization and ferroptosis in cancer immunotherapy.
Methods
Fungal Culturing and Collection
The C. albicans strain was procured from MINGZHOUBIO (B84199, Ningbo, China). The strain was initially cultured in YPD medium and incubated at 28–30 °C for a duration of 24 to 48 hours. Subsequent to incubation, the medium containing C. albicans was centrifuged at 2500 rpm for 15 minutes. The supernatant was then discarded, leaving behind the C. albicans pellet. This pellet was collected and washed with distilled water two to three times to remove any remaining medium components. Ultimately, the C. albicans pellet was dried to yield the dried C. albicans material ready for further use.
β-Glucan Preparation
As previously described,20 C. albicans β-glucan was obtained by dehydrating fungal cell walls with repeated hot alkali and acid extraction. The process started with treating fungal cell walls at 100 °C with a 1% NaOH solution for 24 hours. The insoluble residue, containing β-glucan, was washed until neutral and centrifuged at 3000 rpm for 15 minutes to isolate the alkali-insoluble β-glucan. Next, this residue was treated with 0.5 M acetic acid at 80 °C for 24 hours to extract the acid-insoluble β-glucan. Afterward, the material was washed extensively with distilled water to reach a neutral pH, separating the β-glucan that is insoluble in both alkali and acid. The resulting β-glucan aggregates were freeze-dried and stored at 4 °C.
Animal Treatment
C57BL/6 mice, aged approximately 6–8 weeks, were purchased from Chengdu Dossy experimental animals Co. Ltd. (Chengdu, China) and maintained on a 12 h light/12 h dark cycle at 25 °C for a week. The Committee on the Protection and Use of Animals in the Southwest Jiaotong University approved the experiments. Next, the mice were randomly grouped and subjected to solid tumor induction. Briefly, mice were injected subcutaneously (s. c). with a mixture of 2.5 * 105 TAMs and 106 LLC cells into the right flank.21 Volume of palpable tumors was monitored and was calculated using the equation (length x width2/ 2). In the β-glucan treatment group, TAMs were pre-treated with β-glucan at a concentration of 50 μg/mL for 24 hours. To further explore the impact of ferroptosis inhibition on TAMs following β-glucan stimulation, an additional group of animal models was introduced. In this group, Ferrostatin-1 (Fer-1, S7243, Selleck) was administered intraperitoneally at a dosage of 10 mg/kg/day, starting from the day of subcutaneous tumor cell injection.
Cell Culture and Treatment
The LLC and B16 cell lines were procured from Procell Life Science & Technology Co., Ltd. (Wuhan, China). They were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM, Thermo Fisher Scientific) supplemented with 10% Fetal Bovine Serum (FBS, Gibco) and 1% Penicillin/Streptomycin (Thermo Fisher Scientific). Incubation was maintained at 37°C in an atmosphere containing 5% CO2. For the generation of bone marrow-derived macrophages (BMDM), bone marrow cells were extracted from the tibias and femurs of 6–8 week-old C57BL/6J mice. These cells were then cultured in DMEM supplemented with 20% FBS, 1% Penicillin/Streptomycin, and 20 ng/mL Macrophage Colony-Stimulating Factor (M-CSF, 315–02, Peprotech) for 7 days to differentiate into macrophages.
Conditioned Medium (CM)
The first conditioned medium (CM1) was generated by culturing tumor cells, and the resulting supernatant was utilized for the cultivation of bone marrow-derived macrophages (BMDM). After a 24-hour incubation in CM1, the BMDM were differentiated into tumor-associated macrophages (TAMs). Subsequently, the second conditioned medium (CM2) was prepared by incubating TAMs in a medium that was either supplemented with dimethyl sulfoxide (DMSO, serving as the control), β-glucans at a concentration of 50 μg/mL, or a combination of Fer-1 and β-glucans. Ultimately, the supernatant from the TAM culture was harvested and used to treat LLC and B16 cell lines.
Cell Viability Assay
Cell viability was assessed using the Cell Counting Kit-8 (CCK-8, C0042, Beyotime). LLC cells were plated in 96-well plates at a density of 1 × 105 cells per well, while B16 cells were seeded at 0.8 × 105 cells per well. On the subsequent day, the cells were exposed to the second conditioned medium (CM2) from the control, β-glucan, and Fer-1+β-glucan groups for a duration of 24 hours. Thereafter, 10 µL of CCK-8 reagent was combined with 90 µL of serum-free Dulbecco’s Modified Eagle’s Medium (DMEM) and added to each well. Following incubation at 37°C for 1 to 4 hours, the absorbance at 450 nm was recorded using a microplate reader.
Scratch Test
LLC and B16 cells were plated in 12-well plates at densities of 8 × 105 and 6 × 105 cells per well, respectively. The following day, a scratch wound was introduced to the cell monolayer using a pipette tip. Subsequently, the wells were rinsed with phosphate-buffered saline (PBS) to remove any detached cells and debris. The initial gap was then photographed under a microscope. Subsequently, the cells were treated with the second conditioned medium (CM2) from various experimental groups. After a 24-hour incubation period, the remaining floating cells were removed by washing with PBS, and the scratch distance was reassessed by a microscope.
Western Blot
Cell samples were harvested and lysed using RIPA Lysis Buffer supplemented with 1 mM phenylmethanesulfonyl fluoride (PMSF) and 1 mM protease inhibitor cocktail. Protein concentrations were determined using the BCA protein assay kit. Subsequently, approximately 20 μg of each protein sample was resolved by 12% SDS-PAGE and transferred onto a polyvinylidene fluoride (PVDF) membrane (Millipore, USA). The PVDF membrane was blocked with 5% skim milk in Tris-buffered saline with Tween-20 (TBST, pH 7.5) for 2 hours at room temperature. The primary antibodies, anti-GPX4 (ab125066, abcam), anti-ACSL4 (ab155282, abcam), and anti-β-actin (66009-1-Ig, Proteintech), were applied overnight at 4 °C. After three washes with TBST, the membrane was incubated with the appropriate secondary antibodies for 2 hours at room temperature. Following another three washes with TBST, the membrane was scanned using eBlot Touch Imager.
Flow Cytometry Analysis
TAMs were resuspended in 100 μL of PBS and incubated with PE-Cy™ 7 Rat Anti-Mouse CD11b (BD Biosciences, Franklin Lakes, NJ), PE Rat Anti-Mouse F4/80 (eBioscience, San Diego, CA), Alexa Fluor™ 700 Rat Anti-Mouse CD206 (eBioscience, San Diego, CA) and APC Rat Anti-Mouse CD86 (eBioscience, San Diego, CA) on ice for 30 min. Cells were washed three times with PBS and resuspended in PBS for FACS analysis. All staining processes were performed according to recommended protocols. The stained cells were detected by SONY MA900 flow cytometer (SONY, Tokyo, Japan). FACS data analysis was performed using FLOWJOTM Software.
Quantitative Real-Time PCR
Total RNA was isolated from TAMs using TRIzol reagent (Vazyme, China). The RNA was subsequently reverse transcribed into complementary DNA (cDNA) using HiScript II Reverse Transcriptase (Vazyme, China). Quantitative real-time polymerase chain reaction (qRT-PCR) was conducted on a Real-Time PCR Instrument, employing a 2x TaqSYBR Green qPCR Mix (Vazyme, China). The data were analyzed using the comparative 2−ΔΔCT method, with gene expression levels normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA. The qRT-PCR primer sequences for the murine genes studied were as follows: IL-1β (forward: 5′-TGCCACCTTTTGACAGTGATG-3′, reverse: 5′-AAGGTCCACGGGAAAGACAC-3′), IL-4 (forward: 5′-TCGGCATTTTGAACGAGGTC-3′, reverse: 5′-GAAAAGCCCGAAAGAGTCTC-3′), IL-6 (forward: 5′-CCAAGAGGTGAGTGCTTCCC-3′, reverse: 5′-CTGTTGTTCAGACTCTCTCCCT-3′), IL-10 (forward: 5′-GCTCTTACTGACTGGCATGAG-3′, reverse: 5′-CGCAGCTCTAGGAGCATGTG-3′), iNOS (forward: 5′-GTTCTCAGCCCAACAATACAAGA-3′, reverse: 5′-GTGGACGGGTCGATGTCAC-3′), TNF-α (forward: 5′-CCCTCACACTCAGATCATCTTCT-3′, reverse: 5′-GCTACGACGTGGGCTACAG-3′), Arginase 1 (forward: 5′-ATGCTCACACTGACATCAAC-3′, reverse: 5′-TCTCTTCCATCACCTTGCC-3′), and CD206 (forward: 5′-CTCTGTTCAGCTATTGGACGC-3′, reverse: 5′-CGGAATTTCTGGGATTCAGCTTC-3′).
FerroOrange, Liperfluo and Lipid ROS Staining
TAMs were seeded into 96-well black plates and treated with either dimethyl sulfoxide (DMSO), β-glucan, or a combination of Fer-1 (pre-treated for 2 hours) and β-glucan for a duration of 24 hours. Next, the TAMs cells were incubated with Ferro Orange working solution (F374, Dojindo), Liperfluo working solution (L248, Dojindo) as well as BDP 581/591 C11 working solution (L267, Dojindo) at 37 °C for 30 min, respectively. After three washes with phosphate-buffered saline (PBS), the cells were further incubated with Hoechst stain at 37°C for 15 minutes. The stained cells were then visualized using a laser scanning confocal microscope.
Statistical Analysis
Statistical analysis was conducted using GraphPad Prism version 8.0. Data are presented as the mean ± standard error of the mean (SEM), based on a minimum of three independent biological replicates. For comparisons between two groups, an unpaired Student’s t test was applied, while one-way or two-way analysis of variance (ANOVA) was utilized to assess differences among multiple groups. Statistical significance was defined as a p value less than 0.05.
Results
β-Glucan Extracted from C. Albicans Inhibited Lung Cancer Growth in vivo and in vitro
To assess the impact of β-glucan on TAMs and subsequent tumor growth, both in vivo animal models and in vitro cellular assays were conducted (Figure 1A). Initially, β-glucan-treated TAMs were generated by culturing BMDM with tumor cell supernatant, followed by a 24-hour treatment with β-glucan. In the in vivo experiments, mice were inoculated with β-glucan-treated TAMs along with tumor cells. For the in vitro component, tumor cells were exposed to the supernatant from β-glucan-treated TAMs. In comparison to the control (Ctrl) group, the β-glucan treatment group exhibited a noticeable reduction in tumor size and weight progression (Figure 1B–D). The influence of β-glucan on cellular proliferation and migration was further quantified using Scratch assays and the CCK8 assay. Consistent with the animal findings, the data revealed that the β-glucan group had significantly diminished migratory activity and a reduced rate of cell proliferation compared with those in Ctrl group (Figure 1E–G). These findings suggest that β-glucan extracted from C. albicans possesses the potential to suppress the growth of lung cancer cells.
 |
Figure 1 β-glucan extracted from C. albicans inhibited lung cancer growth in vivo and in vitro. (A). Illustration of the establishment of mouse models treated with β-glucan and the process of cell supernatant transfer for culture. (B and C). Comparison of tumor size and tumor volume in mice treated with β-glucan (50 μg/mL) versus the control group after tumor induction. (D). Statistical analysis of tumor weight in the β-glucan-treated (50 μg/mL) and control groups. (E and F). Evaluation of the migratory capacity of LLC and B16 cells exposed to conditioned media (CM2) from β-glucan-treated (50 μg/mL) and control cells for 24 hours. (G). Measurement of the proliferative capacity of LLC and B16 cells treated with different CM2 for 24 hours. All data presented in this study are reported as the mean ± SEM, representative of at least three independent experiments. *p<0.05, ***p<0.001, ****p<0.0001. |
β-Glucan Exerted Antitumor Effects by Inducing M1 Polarization of TAMs
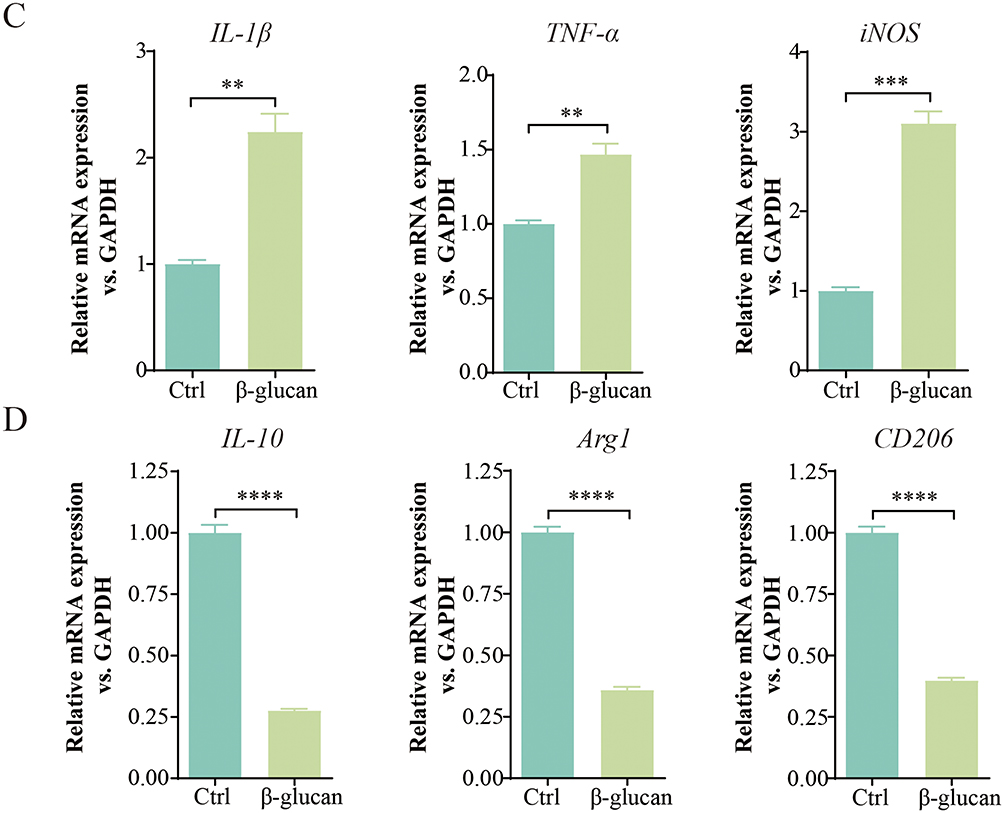
Subsequently, we investigated the phenotype of tumor-associated macrophages (TAMs) to elucidate the specific regulatory mechanisms by which β-glucan influences lung cancer development. Recognizing the immunomodulatory role of TAM polarity in tumor biology, we hypothesized that β-glucan might exert anti-tumor effects by modulating this polarization. Using flow cytometry, we observed that β-glucan treatment led to a significant increase in CD86+ TAMs and a concurrent decrease in CD206+ cells (Figure 2A and B). Furthermore, we quantified the mRNA expression levels of both pro-inflammatory (M1 marker) and anti-inflammatory (M2 marker) cytokines. The results indicated a notable upregulation of IL-1β, TNF-α, and iNOS following β-glucan treatment, while the expression levels of IL-10, Arg1, and CD206 were significantly downregulated (Figure 2C and D). Collectively, these findings suggest that β-glucan facilitates the shift of TAMs towards an M1-like phenotype, potentially enhancing the immune response against lung cancer.
Figure 2 Continued. Figure 2 β-glucan exerted antitumor effects by inducing M1 polarization of TAMs. (A and B). Flow cytometry was utilized to determine the percentages of CD86 and CD206 expression in TAMs following a 24-hour stimulation with β-glucan at a concentration of 50 μg/mL. (C and D). qRT-PCR was employed to measure the mRNA levels of inflammatory markers IL-1β, TNF-α, iNOS, as well as anti-inflammatory markers IL-10, Arg1, and CD206 in TAMs after a 24-hour treatment with β-glucan (50 μg/mL). All data presented in this study are reported as the mean ± SEM, representative of at least three independent experiments. **p<0.01, ***p<0.001, ****p<0.0001.
β-Glucan Activated Ferroptosis in TAMs
To further elucidate the molecular underpinnings of β-glucan-mediated polarization of TAMs, we conducted a comprehensive analysis of RNA sequencing data from bone marrow-derived macrophages (BMDM) following β-glucan treatment, obtained from the Gene Expression Omnibus (GEO) database. Our analysis identified a total of 652 differentially expressed genes (DEGs), comprising 314 up-regulated and 342 down-regulated genes (Figure 3A and Supplementary Table 1). Notably, KEGG pathway analysis revealed a significant enrichment of these DEGs in the ferroptosis pathway, which presented the most significant p-value (Figure 3B). Concurrently, Gene Set Enrichment Analysis (GSEA) substantiated a positive correlation between β-glucan treatment and the induction of ferroptosis (Figure 3C). Expanding on this, we examined key ferroptosis-associated markers. ACSL4, an enzyme pivotal in the metabolism of polyunsaturated fatty acids (PUFAs), is known to initiate ferroptosis upon its upregulation.22 Conversely, GPX4, a glutathione-dependent enzyme that mitigates lipid peroxides by converting them into lipid alcohols, thus preventing ferroptosis.23 In contrast to the control group, the β-glucan treatment group exhibited a significant upregulation of ACSL4 and a concurrent downregulation of GPX4 expression (Figure 3D). Furthermore, employing FerroOrange probes, we quantified intracellular ferrous ion levels in TAMs, observing a marked elevation after a 24-hour β-glucan treatment. This increase in ferrous ions was corroborated by Liperfluo and C11 BODIPY staining, which also indicated an augmentation in lipid peroxides (Figure 3E–J). These observations lead us to hypothesize that ferroptosis potentially plays a crucial role in the polarization of TAMs.
Figure 3 Continued. Figure 3 β-glucan activated ferroptosis in TAMs. (A). The volcano plot illustrates differentially expressed genes (DEGs) in bone marrow-derived macrophages (BMDM) following β-glucan treatment. (B). KEGG pathway analysis depicting the enrichment of DEGs in BMDM treated with β-glucan. (C) Gene Set Enrichment Analysis (GSEA) of the global gene expression in BMDM after β-glucan exposure. (D). Western blot analysis of ACSL4 and GPX4 protein levels in TAMs treated with β-glucan (50 μg/mL) for 24 hours. (E). Intracellular ferrous ion levels in TAMs, measured using FerroOrange probes after a 24-hour β-glucan (50 μg/mL) treatment. (F). Confocal microscopy images showing changes in lipid peroxidation in TAMs following Liperfluo probe staining, post β-glucan treatment. (G). Quantitative assessment of the oxidized to nonoxidized lipid ratio in TAMs, as visualized by confocal microscopy after C11-BODIPY probe staining. (H-J). Statistical analysis of intracellular ferrous ions, lipid peroxidation levels, and the oxidized to nonoxidized lipid ratio. All data presented in this study are reported as the mean ± SEM, representative of at least three independent experiments. *p<0.05, **p<0.01, ****p<0.0001.

β-Glucan Converted the TAMs Polarization to M1-Like Phenotype via the Ferroptosis
Different forms of cell death exert distinct influences on the polarization of macrophage phenotypes.24 To delineate the contribution of ferroptosis to the polarization of TAMs, we utilized a specific inhibitor to examine its impact. TAMs were pre-treated with the ferroptosis inhibitor Fer-1 for 2 hours prior to a 24-hour exposure to β-glucan. Initially, we evaluated the levels of ferrous ion and lipid peroxides following Fer-1 treatment. It showed that Fer-1 effectively reversed the elevations in ferrous ion and lipid peroxides induced by β-glucan (Figure 4A–C). Furthermore, flow cytometry analysis revealed that Fer-1 treatment reduced the population of CD86+ macrophages and increased CD206+ macrophages in comparison to the group treated with β-glucan alone (Figure 5A and B). Additionally, our results indicated that Fer-1 significantly suppressed the expression of pro-inflammatory markers IL-6 and iNOS, while concurrently enhancing the expression of anti-inflammatory cytokines IL-4 and IL-10 (Figure 5C). In a further exploration, we examined the potential influence of apoptosis and pyroptosis on TAM polarization after β-glucan treatment using Z-VAD-FMK and Disulfiram, respectively. Contrary to Fer-1, these inhibitors showed no significant effect on the expression levels of M1 and M2 markers, implying a limited role in the polarization process induced by β-glucan (Figure 5D). These findings collectively underscore the role of β-glucan in modulating TAM polarization through the induction of ferroptosis.
 |
Figure 4 β-glucan converted the TAMs polarization to M1-like phenotype via the ferroptosis. (A). TAMs were pre-treated with the ferroptosis inhibitor Fer-1 (10 μM) for 2 hours prior to a 24-hour treatment with β-glucan (50 μg/mL). Representative images of intracellular ferrous ions were captured using FerroOrange probes. (B). Confocal microscopy was employed to visualize alterations in lipid peroxidation within TAMs after staining with Liperfluo probes. (C). The ratio of oxidized to nonoxidized lipids in TAMs was quantified using confocal microscopy following staining with C11-BODIPY probes. All data presented in this study are reported as the mean ± SEM, representative of at least three independent experiments. ***p<0.001, ****p<0.0001. |
 |
Figure 5 Continued. |
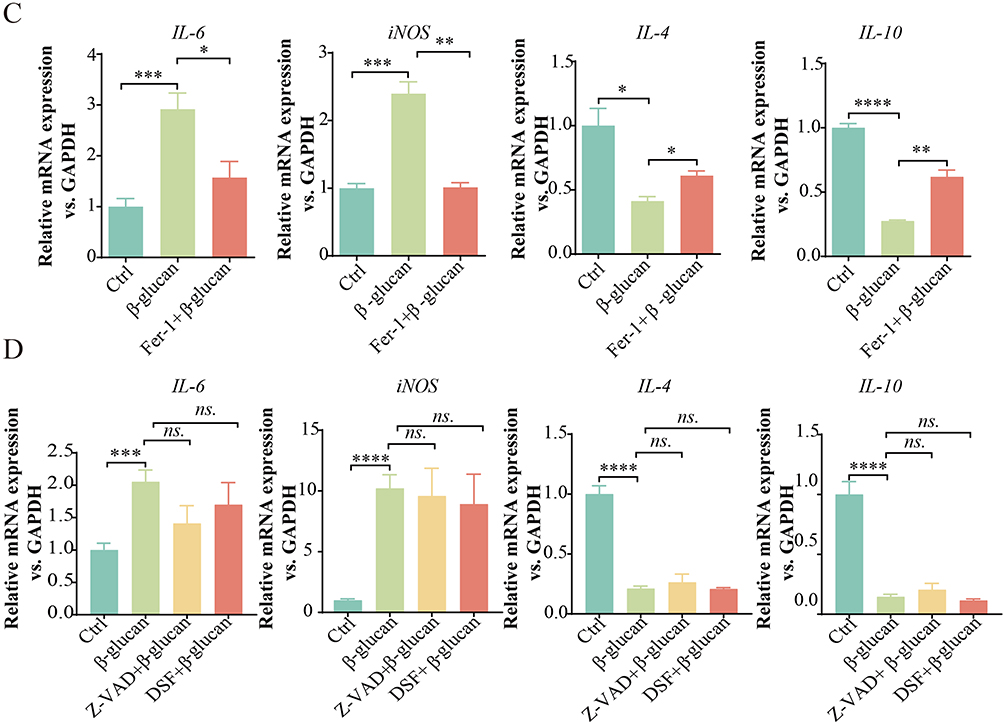 |
Figure 5 β-glucan converted the TAMs polarization to M1-like phenotype via the ferroptosis. (A and B). TAMs were pre-treated with the ferroptosis inhibitor Fer-1 (10 μM) for 2 hours prior to a 24-hour treatment with β-glucan (50 μg/mL). Flow cytometry was utilized to determine the percentages of CD86+ macrophages and CD206+ macrophages. (C and D). qRT-PCR was employed to measure the mRNA levels of IL-6, iNOS, IL-4 and IL-10 in TAMs in each group. All data presented in this study are reported as the mean ± SEM, representative of at least three independent experiments. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001. ns.: Not significant. |
Ferroptosis Inhibitor Reversed the Tumor-Suppressive Effects of β-Glucan Both in vivo and in vitro
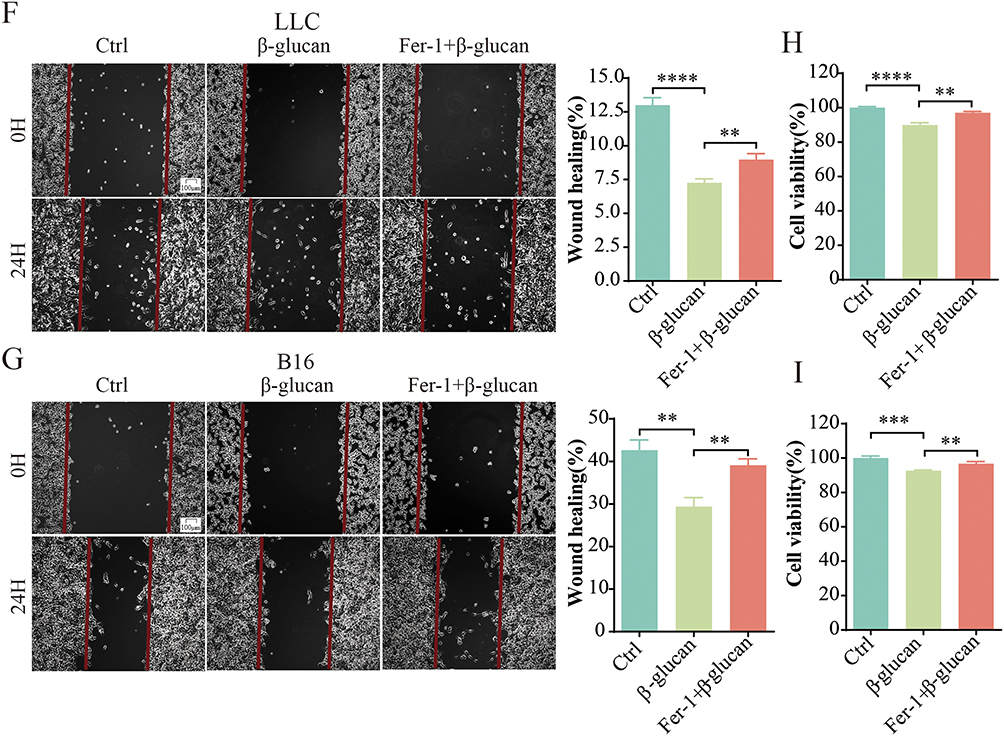
The above results indicated that β-glucan triggered ferroptosis in tumor-associated macrophages (TAMs), which results in an M1-like polarization and consequently inhibits the growth of lung cancer. Building on these insights, we extended our investigation to assess the efficacy of Fer-1 in negating the tumor-suppressive effects triggered by β-glucan both in vitro and in vivo (Figure 6A and E). In the in vivo experiments, co-treatment with Fer-1 and β-glucan resulted in a significant increase in tumor size and weight relative to treatment with β-glucan alone (Figure 6B–D). Paralleling these observations, in vitro analysis of the lung cancer cell lines LLC and B16 revealed that the CM of combination of Fer-1 and β-glucan led to higher rates of cell proliferation and migration compared to the CM of β-glucan treatment alone (Figure 6F–I). The collective data demonstrated that Fer-1 can indeed reverse the growth-suppressive impact of β-glucan derived from C. albicans on lung cancer cells.
Figure 6 Continued. Figure 6 Ferroptosis inhibitor reversed the tumor-suppressive effects of β-Glucan both in vivo and in vitro. (A). Schematic representation illustrating the establishment of mouse models treated with Fer-1 in combination with β-glucan. (B and C). Comparison of tumor size and tumor volume among groups treated with Fer-1 (10 μM) plus β-glucan, β-glucan alone (50 μg/mL), and the control group after tumor induction. (D). Statistical evaluation of tumor weight in the Fer-1 (10 μM) plus β-glucan treated group, β-glucan treated group (50 μg/mL), and the control group. (E). Schematic representation illustrating the establishment of cell models treated with Fer-1 in combination with β-glucan. (F and G). Assessment of the migratory capacity of LLC and B16 cells exposed to conditioned media (CM2) from the Fer-1 (10 μM) plus β-glucan treated group, β-glucan treated group (50 μg/mL), and the control group for 24 hours, using a scratch test. (H and I). Measurement of the proliferative capacity of LLC and B16 cells following treatment with CM2 from the Fer-1 (10 μM) plus β-glucan treated group, β-glucan treated group (50 μg/mL), and the control group for 24 hours, as determined by the CCK8 assay. All data presented in this study are reported as the mean ± SEM, representative of at least three independent experiments. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001.
Discussion
The findings of this study provide compelling evidence that β-glucan extracted from C. albicans exerts significant inhibitory effects on lung cancer growth, both in vivo and in vitro. Our results underscore the potential of β-glucan as a therapeutic agent, highlighting its multifaceted role in modulating the TME and the polarization of TAMs.
Previous research has documented the anti-metastatic effects of β-glucan derived from Lentinus edodes, particularly its ability to suppress the spread of breast tumors to the lungs.25 Additionally, studies have shown that yeast-derived particulate β-glucan can diminish tumor growth in mice bearing tumors.26 Aligning with these findings, our current investigation, encompassing both in vivo and in vitro assays, has consistently demonstrated that β-glucan treatment results in a significant reduction in tumor size, weight, and cellular proliferation. These results reinforce the direct suppressive effect of β-glucan on lung cancer cells. Our preliminary conclusions suggest that β-glucan from C. albicans not only inhibits the growth of lung tumor cells but also serves as a preventative measure against the advancement of lung cancer.
Macrophage polarization plays a key role in lung cancer progression. One of the traits of malignant tumor is the polarization of TAMs from a pro‐immune (M1‐like) to an immunosuppressive (M2‐like) phenotype.27 For instance, MSC-EV has been shown to enhance miR-21-5p transmission following hypoxia pre-challenge, thereby promoting lung cancer development through reduced apoptosis and induced M2 macrophage polarization.28 Additionally, tumoral NOX4 is known to recruit M2 TAMs, further promoting the growth of non-small cell lung cancer.29 Several studies have reported that β-glucan treatment inhibited the expression of M2-type markers, while increased the expression of M1-type markers,30 and converted the immunosuppressive M2 and TAMs into M1 anti-tumor phenotype.31–33 In this study, both flow cytometry and real-time fluorescence analysis confirmed that β-glucan reprogrammed TAMs from an M2- to an M1-dominant phenotype. β-glucan treatment facilitated M1 polarization of macrophages, particularly in the presence of lung tumor cell supernatant. Concurrently, M1 macrophages exhibited antitumor and pro-inflammatory effects, which were contingent on the secretions of tumor cells in the presence of β-glucan. Therefore, we propose that β-glucan mediates its inhibitory effect on lung cancer by promoting the transformation of TAMs into an M1-like phenotype, thereby enhancing the immune response against the tumor.
Ferroptosis, a recently characterized form of regulated cell death, is distinguished by the accumulation of iron-dependent lipid peroxidation. It has been reported that the inhibition of APOC1 has been shown to drive the transition of M2 macrophages to the M1 phenotype through the ferroptosis pathway, thereby inhibiting the growth of hepatocellular carcinoma.34 Additionally, dihydroartemisinin has demonstrated anti-tumor effects in lung cancer by inducing macrophage reprogramming to an M1 phenotype via ferroptosis-mediated DNA damage.35 However, the connection between β-glucan-induced macrophage polarization and ferroptosis has not been previously explored. In this study, we investigated the potential for β-glucan to regulate macrophage polarization through the induction of ferroptosis. The RNA sequencing analysis indicated a link between ferroptosis and β-glucan treatment in bone marrow-derived macrophages (BMDM). Subsequent Western blotting confirmed that β-glucan significantly altered the protein levels of ACSL4 and GPX4 in TAMs, both of which are key players in promoting ferroptosis. Finally, immunofluorescence assays revealed an increase in intracellular ferrous ion levels and lipid peroxidation in TAMs, which were attenuated by pre-treatment with the ferroptosis inhibitor Fer-1. These findings collectively illuminate the mechanism by which β-glucan reprograms macrophage polarization, potentially through the activation of ferroptosis pathways.
Ferrostatin-1 (Fer-1), a well-established inhibitor of ferroptosis, has been shown to augment the protective effects of uridine, thereby inhibiting macrophage ferroptosis and offering therapeutic benefits against lipopolysaccharide (LPS)-induced acute lung injury (ALI).36 Furthermore, Fer-1 has been demonstrated to inhibit Tim-AIII-mediated lung cancer cell death, promoting tumor progression.37 Our study presents data indicating that pre-treatment with Fer-1 mitigated the M1 polarization of macrophages induced by β-glucan, highlighting the contribution of ferroptosis to this polarization in tumor-associated macrophages (TAMs). In contrast, pre-treatment with inhibitors of apoptosis or pyroptosis did not impact the markers of M1 and M2 macrophages, suggesting that these forms of cell death are not predominant in β-glucan-induced cell death. Additionally, our findings revealed that Fer-1 pre-treatment significantly enhanced tumor size and weight in murine models, and the supernatant from tumor-associated macrophages (TMAs) stimulated tumor cell proliferation and migration in vitro, as compared to the β-glucan-only group.
In conclusion, β-glucan from C. albicans emerges as a promising candidate for immunotherapy in lung cancer, with its effects potentially extending beyond direct cytotoxicity to include the modulation of macrophage polarization and the induction of ferroptosis (Figure 7). These findings open new avenues for research into the development of novel therapeutics that harness the immunomodulatory potential of β-glucan and its impact on ferroptosis in cancer treatment.
 |
Figure 7 A diagram of possible mechanism by which β-glucan extracted from Candida albicans inhibits the development of lung cancer. β-glucan extracted from C. albicans can promote the ferroptosis of TAMs, resulting in increased secretion of pro-inflammatory cytokines and reduced secretion of anti-inflammatory cytokines. This process promotes the polarization of TAMs to M1 macrophages, ultimately inhibiting the progression of lung cancer. |
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (82370022, 82370023, and 82300088), China Postdoctoral Science Foundation (2023M730799), Natural Science Foundation of Sichuan Province (23NSFSC1736, 24NSFSC2826), Science and Technology Project of Health Commission of Sichuan Province (24QNMP022), Chengdu High-level Key Clinical Specialty Construction Project (ZX20201202020).
Disclosure
The authors declare that they have no conflict of interest.
References
1. Thai AA, Solomon BJ, Sequist LV, et al. Lung cancer. Lancet. 2021;398(10299):535–554. doi:10.1016/S0140-6736(21)00312-3
2. Health Commission of the PRC N, Health Commission Of The People’s Republic Of China.. National guidelines for diagnosis and treatment of lung cancer 2022 in China (English version). Chin J Cancer Res. 2022;34(3):176–206. doi:10.21147/j.issn.1000-9604.2022.03.03
3. Mok TSK, Wu Y-L, Kudaba I, et al. Pembrolizumab versus chemotherapy for previously untreated, PD-L1-expressing, locally advanced or metastatic non-small-cell lung cancer (KEYNOTE-042): a randomised, open-label, controlled, Phase 3 trial. Lancet. 2019;393(10183):1819–1830. doi:10.1016/S0140-6736(18)32409-7
4. Bai R, Li Y, Jian L, Yang Y, Zhao L, Wei M. The hypoxia-driven crosstalk between tumor and tumor-associated macrophages: mechanisms and clinical treatment strategies. Mol Cancer. 2022;21(1):177.
5. Campbell MJ, Tonlaar NY, Garwood ER, et al. Proliferating macrophages associated with high grade, hormone receptor negative breast cancer and poor clinical outcome. Breast Cancer Res Treat. 2011;128(3):703–711. doi:10.1007/s10549-010-1154-y
6. Chung FT, Lee K-Y, Wang C-W, et al. Tumor-associated macrophages correlate with response to epidermal growth factor receptor-tyrosine kinase inhibitors in advanced non-small cell lung cancer. Int J Cancer. 2012;131(3):E227–35. doi:10.1002/ijc.27403
7. Conway EM, Pikor LA, Kung SHY, et al. Macrophages, Inflammation, and Lung Cancer. Am J Respir Crit Care Med. 2016;193(2):116–130. doi:10.1164/rccm.201508-1545CI
8. Mantovani A, Sozzani S, Locati M, et al. Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002;23(11):549–555. doi:10.1016/S1471-4906(02)02302-5
9. Yang L, Zhang Y. Tumor-associated macrophages, potential targets for cancer treatment. Biomark Res. 2017;5(1):25. doi:10.1186/s40364-017-0106-7
10. Shi L, Liu Y, Li M, et al. Emerging roles of ferroptosis in the tumor immune landscape: from danger signals to anti-tumor immunity. FEBS J. 2022;289(13):3655–3665. doi:10.1111/febs.16034
11. Zhou Z, Xu B, Hu N, et al. Targeting the Macrophage-Ferroptosis Crosstalk: a Novel Insight into Tumor Immunotherapy. Front Biosci. 2022;27(7):203. doi:10.31083/j.fbl2707203
12. Chen X, Kang R, Kroemer G, et al. Broadening horizons: the role of ferroptosis in cancer. Nat Rev Clin Oncol. 2021;18(5):280–296. doi:10.1038/s41571-020-00462-0
13. Dai E, Han L, Liu J, et al. Autophagy-dependent ferroptosis drives tumor-associated macrophage polarization via release and uptake of oncogenic KRAS protein. Autophagy. 2020;16(11):2069–2083. doi:10.1080/15548627.2020.1714209
14. Hu Z-W, Wen Y-H, Ma R-Q, et al. Ferroptosis driver SOCS1 and suppressor FTH1 independently correlate with M1 and M2 macrophage infiltration in head and neck squamous cell carcinoma. Front Cell Dev Biol. 2021;9:727762. doi:10.3389/fcell.2021.727762
15. Ruiz-Herrera J, Elorza MV, Valentín E, et al. Molecular organization of the cell wall of Candida albicans and its relation to pathogenicity. FEMS Yeast Res. 2006;6(1):14–29. doi:10.1111/j.1567-1364.2005.00017.x
16. Garcia-Rubio R, de Oliveira HC, Rivera J, et al. The fungal cell wall: Candida, Cryptococcus, and Aspergillus species. Front Microbiol. 2019;10:2993. doi:10.3389/fmicb.2019.02993
17. Sadeghi F, Peymaeei F, Falahati M, et al. The effect of Candida cell wall beta-glucan on treatment-resistant LL/2 cancer cell line: in vitro evaluation. Mol Biol Rep. 2020;47(5):3653–3661. doi:10.1007/s11033-020-05459-7
18. Stothers CL, Burelbach KR, Owen AM, et al. beta-glucan induces distinct and protective innate immune memory in differentiated macrophages. J Immunol. 2021;207(11):2785–2798. doi:10.4049/jimmunol.2100107
19. Wang Q, Zhang ZF, Zheng GH, et al. beta-Glucans: relationships between modification, conformation and functional activities. Molecules. 2017;22(2):1315.
20. Nisini R, Torosantucci A, Romagnoli G, et al. β-Glucan of Candida albicans cell wall causes the subversion of human monocyte differentiation into dendritic cells. J Leukoc Biol. 2007;82(5):1136–1142. doi:10.1189/jlb.0307160
21. Yin H, Zhang X, Yang P, et al. RNA m6A methylation orchestrates cancer growth and metastasis via macrophage reprogramming. Nat Commun. 2021;12(1):1394. doi:10.1038/s41467-021-21514-8
22. Chen X, Li J, Kang R, et al. Ferroptosis: machinery and regulation. Autophagy. 2021;17(9):2054–2081. doi:10.1080/15548627.2020.1810918
23. Yan HF, Zou T, Tuo Q-Z, et al. Ferroptosis: mechanisms and links with diseases. Signal Transduct Target Ther. 2021;6(1):49. doi:10.1038/s41392-020-00428-9
24. Shao Y, Wang Y, Su R, et al. Dual identity of tumor-associated macrophage in regulated cell death and oncotherapy. Heliyon. 2023;9(7):e17582. doi:10.1016/j.heliyon.2023.e17582
25. Zhu F, Zhang Q, Feng J, et al. beta-Glucan produced by Lentinus edodes suppresses breast cancer progression via the inhibition of macrophage M2 polarization by integrating autophagy and inflammatory signals. Immun Inflamm Dis. 2023;11(5):e876. doi:10.1002/iid3.876
26. Albeituni SH, Ding C, Liu M, et al. Yeast-derived particulate beta-glucan treatment subverts the suppression of myeloid-derived suppressor cells (MDSC) by inducing polymorphonuclear MDSC apoptosis and monocytic MDSC differentiation to APC in cancer. J Immunol. 2016;196(5):2167–2180. doi:10.4049/jimmunol.1501853
27. Yang J, Zhang Z, Chen C, et al. MicroRNA-19a-3p inhibits breast cancer progression and metastasis by inducing macrophage polarization through downregulated expression of Fra-1 proto-oncogene. Oncogene. 2014;33(23):3014–3023. doi:10.1038/onc.2013.258
28. Ren W, Hou J, Yang C, et al. Extracellular vesicles secreted by hypoxia pre-challenged mesenchymal stem cells promote non-small cell lung cancer cell growth and mobility as well as macrophage M2 polarization via miR-21-5p delivery. J Exp Clin Cancer Res. 2019;38(1):62. doi:10.1186/s13046-019-1027-0
29. Zhang J, Li H, Wu Q, et al. Tumoral NOX4 recruits M2 tumor-associated macrophages via ROS/PI3K signaling-dependent various cytokine production to promote NSCLC growth. Redox Biol. 2019;22:101116. doi:10.1016/j.redox.2019.101116
30. Cheng H, Sun L, Shen D, et al. Beta-1,6 glucan converts tumor-associated macrophages into an M1-like phenotype. Carbohydr Polym. 2020;247:116715. doi:10.1016/j.carbpol.2020.116715
31. Sun J, Sun J, Song B, et al. Fucoidan inhibits CCL22 production through NF-kappaB pathway in M2 macrophages: a potential therapeutic strategy for cancer. Sci Rep. 2016;6(1):35855. doi:10.1038/srep35855
32. Zhang M, Wu W, Ren Y, et al. Structural characterization of a novel polysaccharide from Lepidium meyenii (Maca) and analysis of its regulatory function in macrophage polarization in vitro. J Agric Food Chem. 2017;65(6):1146–1157. doi:10.1021/acs.jafc.6b05218
33. Liu Y, Zhang L, Zhu X, et al. Polysaccharide Agaricus blazei Murill stimulates myeloid derived suppressor cell differentiation from M2 to M1 type, which mediates inhibition of tumour immune-evasion via the Toll-like receptor 2 pathway. Immunology. 2015;146(3):379–391. doi:10.1111/imm.12508
34. Hao X, Zheng Z, Liu H, et al. Inhibition of APOC1 promotes the transformation of M2 into M1 macrophages via the ferroptosis pathway and enhances anti-PD1 immunotherapy in hepatocellular carcinoma based on single-cell RNA sequencing. Redox Biol. 2022;56:102463. doi:10.1016/j.redox.2022.102463
35. Li LG, Peng X-C, Yu -T-T, et al. Dihydroartemisinin remodels macrophage into an M1 phenotype via ferroptosis-mediated DNA damage. Front Pharmacol. 2022;13:949835. doi:10.3389/fphar.2022.949835
36. Lai K, Song C, Gao M, et al. Uridine alleviates sepsis-induced acute lung injury by inhibiting ferroptosis of macrophage. Int J Mol Sci. 2023;24(6):5093. doi:10.3390/ijms24065093
37. Zhou C, Yu T, Zhu R, et al. Timosaponin AIII promotes non-small-cell lung cancer ferroptosis through targeting and facilitating HSP90 mediated GPX4 ubiquitination and degradation. Int J Biol Sci. 2023;19(5):1471–1489. doi:10.7150/ijbs.77979
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.