")
Back to Journals » Vascular Health and Risk Management » Volume 21
Causal Relationship Between Immune Cells and Venous Thromboembolism: A Bidirectional Two-Sample Mendelian Randomization Study
Authors Su Q , Li Y, Wen C, Li L, Ye Q, Chen M, Xie L, Hu C, Wu H
Received 23 September 2024
Accepted for publication 7 March 2025
Published 25 March 2025 Volume 2025:21 Pages 181—195
DOI https://doi.org/10.2147/VHRM.S497476
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Akash Batta
Qiwen Su,1,2 Yue Li,1,2 Cheng Wen,1,2 Lilong Li,1,2 Qianling Ye,2 Ming Chen,2 Linyang Xie,1,2 Chenming Hu,1,2 Huaping Wu1,2
1School of Clinical Medicine, North Sichuan Medical College, Nanchong, People’s Republic of China; 2Department of Vascular Surgery, Dazhou Central Hospital, Dazhou, People’s Republic of China
Correspondence: Huaping Wu, Department of Vascular Surgery, Dazhou Central Hospital, Dazhou, Sichuan, 635000, People’s Republic of China, Email [email protected] Chenming Hu, Department of Vascular Surgery, Dazhou Central Hospital, Dazhou, People’s Republic of China, Email [email protected]
Introduction: Manny evidence indicates that numerous immune cells are linked to the onset and progression of VTE, though the causal relationship remains unclear. To determine the association between immune cells and VTE, we performed a bidirectional two-sample Mendelian randomization (MR) study.
Methods: A comprehensive MR analysis was conducted to ascertain the causal relationship between immune cell signatures and VTE. Leveraging publicly available genetic data, we examined the causal associations between 731 immune cell signatures and the risk of VTE. The analysis encompassed four types of immune signatures, namely median fluorescence intensities, relative cell counts, absolute cell counts, and morphological parameters. We employed the two-sample MR analysis, used the inverse variance-weighted (IVW) approach as the primary analytical method. Rigorous sensitivity analyses were employed to validate the robustness, heterogeneity, and presence of horizontal pleiotropy in the results. Furthermore, the reverse MR analysis was implemented to confirm the existence of reverse causal relationships.
Results: Eighteen immune cell signatures were found to have nominally significant associations with VTE according to the IVW method. The level of CD14 expression on CD14+ CD16+ monocytes (OR 0.95) and ten other phenotypes were identified as protective factors against VTE. Conversely, the percentage of HLA DR+ T cells among lymphocytes (OR 1.03) and six other phenotypes were identified as risk factors associated with an increased likelihood of VTE. The expression level of CX3CR1 on CD14- CD16+ monocytes showed a potential bidirectional causal relationship.
Conclusion: Our study identified 18 types of immune cell signatures that could impact VTE development, offering novel insights for future mechanistic and clinical studies in this field. Further studies to prospectively validate our findings are needed.
Keywords: causal inference, genome-wide association study, immunity, Mendelian randomization analysis, venous thromboembolism
Introduction
Venous thromboembolism (VTE), which includes deep vein thrombosis and pulmonary embolism, is the third most prevalent vascular disease and third leading cause of death worldwide, affecting approximately 10 million people annually.1,2
Over recent years, the incidence of VTE has shown a steady increase along with the increase in VTE risk factors, such as aging populations, obesity, heart failure, and cancer.
VTE is strongly associated with a poor prognosis and compromised quality of life;2 20% of patients with pulmonary embolism die either before diagnosis or shortly thereafter,3 30% of patients with VTE experience recurrence within a 10-year timeframe,4 and 20–50% of patients with deep vein thrombosis experience post-thrombotic syndrome, leading to diminished quality of life and potential disability.5 Moreover, VTE imposes a considerable annual economic burden, estimated at €1.5–3.3 billion in Europe and $7–10 billion in the United States.6,7
While Virchow’s triad of factors, namely, abnormalities in blood flow, blood components, and vessel walls, are the cornerstone of VTE-related pathophysiology, the underlying pathophysiological processes that lead to thrombosis are not fully understood, leaving a substantial gap in terms of VTE prevention and treatment. Inflammation and immune cells are key contributors to the pathogenesis of thrombosis, but the exact mechanisms remain elusive in human populations, and further elucidating their role may contribute to an enhanced understanding of the pathogenesis of VTE and advance prevention and treatment strategies. Current data have been primarily obtained from experimental models, and the relationship between immune cells and VTE in humans is not well established. Therefore, conducting a genetic-level study is pertinent. At present, a large amount of evidence has indicated the role of the innate immune system in the pathogenesis of VTE. However, we still do not understand the exact mechanisms involved.8,9 Khorana AA discussed the complexity of immune responses in cancer-associated VTE, pointing out that while there is significant immune activation, the direct causal pathways between immune cells and thrombosis are not well-defined in human populations Therefore, conducting a genetic-level study is pertinent.10 Mendelian randomization (MR) is a genetic epidemiological method that utilizes single nucleotide polymorphisms (SNPs) as instrumental variables (IVs) to infer the causal relationship between exposure and outcomes.11 This method can be viewed as a “natural” randomized controlled trial because alleles are assigned randomly during meiosis. MR may reduce both traditional confounding variables and reverse causation, thereby offering more robust evidence of causal inference.12 Therefore, we conducted this bidirectional two-sample MR analysis of summary Genome-Wide Association Studies (GWAS) data to evaluate the causal relationship between VTE and immune cells. We aimed to provide insights for the development of new research directions and treatment strategies for VTE. MR by using genetic variants as proxies for immune cell traits, MR can avoid the confounding and reverse causality issues that plague traditional observational studies. This approach is particularly important given the complex, multi-dimensional nature of immune responses, where immune cells might influence VTE through several interconnected pathways, making it difficult to establish direct causality without the proper genetic tools.
Methods
Study Design
MR leverages genetic variations as proxies for risk factors, necessitating the use of valid IVs that adhere to three crucial assumptions for reliable causal inference: (1) direct association of genetic variation with exposure, (2) absence of association between genetic variation and potential confounders in the relationship between exposure and outcome, and (3) assurance that genetic variation does not exert influence on the outcome through pathways unrelated to the exposure.13
We performed an extensive assessment of the causal relationship between 731 immune cell signatures, categorized into seven groups, and VTE using bidirectional two-sample MR analysis. The timeframe from the FinnGen study were collected and analyzed from 2015 to 2022, while the immune cell trait data were extracted from the GWAS Catalog in 2023. The comprehensive design is presented in Figure 1.
 |
Figure 1 A detailed overview of the Mendelian randomization design. Abbreviations: GWAS, Genome-Wide Association Studies; MR, Mendelian randomization; SNP, Single nucleotide polymorphism. |
Study Setting
This study utilized publicly available summary-level genetic data from the FinnGen Study and the GWAS Catalog. The FinnGen data is derived from a large cohort of European individuals, with cases of VTE identified using International Classification of Diseases codes. The analyses were conducted using these publicly accessible datasets.
Study Population-The study population for this MR analysis includes 19,372 patients and 357,905 controls from the FinnGen study, primarily consisting of individuals of European descent. The immune cell trait data were derived from a separate GWAS involving 3757 European individuals.
Ethical Approval
According to the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects (China), as the data used in this study were de-identified and publicly available, ethical approval was not required. The studies incorporated into our analysis received approval from all the respective institutional review boards, and all participants in these studies granted informed consent.
Sample Size
This study utilized data from large-scale GWAS cohorts, so no sample size calculation was required. The large sample sizes of these datasets provide sufficient statistical power to detect causal associations between immune cell signatures and VTE.
Data Sources
GWAS Data Sources for VTE
The most recent FinnGen GWAS results were the source of summary-level statistical data for VTE (available at https://r9.finngen.fi/).14 Venous thromboembolism (VTE) is defined as the obstruction of a vein or veins (embolism) by a blood clot (thrombus) in the bloodstream. It also refers to the occlusion of the lumen of a vein by a thrombus that has migrated from a distal site via the bloodstream. Phenotypic codes for VTE cases were labeled as “I9_VTE”. The analysis, adjusted for age, sex, genetic relatedness, genotyping batch, and the first 10 principal components, involved 19,372 patients and 357,905 controls.
Immunity-Wide GWAS Data Sources
Publicly accessible summary statistics for each immune trait in the GWAS data can be found in the GWAS catalog, spanning the accession numbers GCST0001391 to GCST0002121.15 A comprehensive set of 731 immunophenotypes were examined, consisting of absolute cell counts (n = 118), median fluorescence intensities indicative of surface antigen levels (n = 389), morphological parameters (n = 32), and relative cell counts (n = 192). Specifically, the median fluorescence intensities, absolute cell counts, and relative cell counts encompassed B cells, conventional dendritic cells (cDCs), mature T cells, monocytes, myeloid cells, TBNK (T cells, B cells, and natural killer [NK] cells), and Treg panels. The morphological parameters involved cDCs and TBNK panels. The initial GWAS on immune traits utilized data from 3757 European individuals, ensuring non-overlapping cohorts. Imputation of almost 22 million genotyped SNPs was performed using high-density arrays and a Sardinian sequence-based reference panel. Associations were assessed following adjustments for covariates such as sex and age.16
Reverse MR
The data source for reverse MR was identical to that of the forward MR. In this case, we considered VTE as the exposure and extracted SNPs closely related to VTE (p < 10−8). A process akin to forward MR was employed, involving the removal of linkage disequilibrium (LD) and IVs with an F-statistic < 10. The genera identified as significant in the forward MR analysis were utilized as the outcome, and we conducted a two-sample MR analysis to elucidate the causal relationship between VTE and immune cells.
Selection of IVs
We evaluated 731 immunophenotypes using MR analysis. We identified SNPs related to immune cell signatures and utilized them as IVs. To guarantee the authenticity and precision of the inferred causality between immune cell composition and VTE, we employed the following rigorous criteria to select our IVs: (i) SNPs with phenotypic associations were prioritized at p < 1×10−5. This threshold was selected because GWAS-identified loci for immune cells seldom attain the conventional genome-wide significance level (p < 5×10−8), allowing the inclusion of a broader spectrum of genetic variations potentially relevant to immunophenotypes.17 (ii) The reference panel for our study consisted of European samples of the 1000 Genomes Project; therefore, we computed the LD among the identified SNPs. SNPs with R2 <0.001 within a clumping window of 10,000 kb that exhibited the smallest p-values were retained. (iii) SNPs presenting a minor allele frequency of ≤ 0.01 were systematically discarded. In cases where palindromic SNPs were detected, we inferred the alleles on the forward strand based on allele frequency data. The robustness of each SNP was evaluated using the F-statistic: F = R2/(1-R2) × (N-K-1)/K, where R² indicates the fraction of the variance in the exposure accounted for by the genetic variants, N represents the sample size, and K denotes the number of IVs. Instruments were deemed strong with an F-statistic ≥ 10.18,19 Conversely, those with F-statistics < 10 were categorized as weak instruments and excluded from the analyses.20 Additionally, to ensure the quality of our selected SNPs, we utilized the webpage (http://www.phenoscanner.medschl.cam.ac.uk) to authenticate the extracted SNPs and eliminate potential confounding SNPs that may have a causal relationship with VTE.21
Statistical Analysis
MR Analysis
In this study, we sought to elucidate the causal relationship between immune cells and VTE utilizing MR methodologies, namely, an inverse-variance weighted (IVW) test, weighted mode, MR-Egger regression, weighted median estimator, and simple mode. When dealing with multiple IVs, most of our results were shaped using the IVW method.13,22,23 To account for multiple testing, the false discovery rate (FDR) method was applied to adjust the p-values. To evaluate the heterogeneity and potential outliers in our initial observations, we utilized scatter plots, funnel plots, and leave-one-out plots. Additional methodologies provided depth and breadth, ensuring a comprehensive evaluation of the hypotheses.
Sensitivity Analysis
Considering biases may be involved when using an IVW test, a sensitivity analysis, encompassing tests for heterogeneity, pleiotropy, and leave-one-out sensitivity, was conducted to evaluate the reliability and stability of the MR results. Within the framework of two-sample MR, we addressed instrument heterogeneity using Cochran’s Q statistics. A significant Q statistic emerging either from a value surpassing the count of instruments reduced by one or a p-value < 0.05 signaled potential instrument heterogeneity. Addressing such heterogeneity is essential, as it ensures the reliability of MR estimates and prevents potential biases. We utilized the MR-PRESSO method to identify horizontal pleiotropy, which is a potential bias in MR studies.24,25 This approach juxtaposes the observed distances of SNPs to the regression line with the distances anticipated under a null hypothesis devoid of horizontal pleiotropy. When data indicated horizontal pleiotropy, outlier-corrected estimates were provided. To verify the validity of our conclusions, a leave-one-out sensitivity analysis was performed. This enabled us to determine whether a specific SNP was disproportionately influencing the causal interpretation.26,27
All computational tasks were conducted using R software (version 4.3.1). MR analyses were supported and streamlined using the Two-Sample MR package software (version 0.5.7).
Results
IV Selection and Characteristics
Based on IV-related selection criteria, a comprehensive set of 14,456 SNPs were used as IVs for 731 distinct immunophenotypes. An elaborate breakdown of these chosen IVs can be found in the Supplementary Table 1.
Identification of Potential Causal Immunophenotypes
In our exploration of the causal impact of VTE on immunophenotypes, we utilized a two-sample MR analysis, with the IVW method as the primary analytical tool. Following multiple test adjustments through the false discovery rate (FDR) method, no immune traits reached significance at a level of 0.05. These included B cells, cDCs, maturation stages of T cells, monocytes, myeloid cells, and TBNK cells. In total, 308 SNPs were designated as significant IVs for these genera and are listed in Supplementary Table 2. A comprehensive epidemiological study, using an exhaustive literature survey, identified specific SNPs as potential confounding factors, such as SNP rs115307086 for cancer, SNP rs35583090 for hematological neoplasms, SNP rs10155782 for a previous history of thrombosis, SNP rs111747460 for trauma or immobility, and SNP rs148381451 for hemorrhage. All confounding SNP information is shown in Supplementary Table 3.
Main MR Results
In accordance with the STROBE-MR guidelines, the results are presented as odds ratios (OR) with corresponding 95% confidence intervals (CI). A cumulative assessment of these findings is presented in Figure 2 and Supplementary Table 4. Eighteen immune cell types exhibited significant associations, and the definitive MR results are listed in Supplementary Table 5. Based on IVW analyses, the level of CD14 expression on CD14+ CD16+ monocytes ([OR] 0.95, 95% [CI] 0.906–0.992; p = 0.022), the level of CD19 expression on IgD- CD38+ B cells (OR 0.95, 95% CI 0.914–0.988; p = 0.011), the level of CD45RA expression on terminally differentiated CD8+ T cells (OR 0.96, 95% CI 0.915–0.999; p = 0.044), the level of FSC-A expression on human leukocyte antigen (HLA) DR+ NK cells (OR 0.96, 95% CI 0.942–0.986; p = 0.001), the percentage of CD62L- plasmacytoid dendritic cells among dendritic cells (OR 0.97, 95% CI 0.942–0.989, p = 0.004), the percentage of CD4-CD8- NK T cells among lymphocytes (OR 0.97, 95% CI 0.933–0.999; p = 0.043), the level of FSC-A expression on granulocytes (OR 0.97, 95% CI 0.948–0.994; p = 0.016), the level of herpes virus entry mediator (HVEM) expression on naive CD4+ T cells (OR 0.97, 95% CI 0.950–0.994; p = 0.013), the level of CD40 expression on CD14+ CD16- monocytes (OR 0.97, 95% CI 0.948–0.999; p = 0.040), the percentage of effector memory CD8+ T cells among T cells (OR 0.97, 95% CI 0.952–0.997; p = 0.024), and the absolute count of memory B cells (OR 0.98, 95% CI 0.960–0.992; p = 0.003) were found to act as protective factors against VTE. Conversely, the percentage of HLA DR+ T cells among lymphocytes (OR 1.03, 95% CI 1.006–1.048; p = 0.013), the percentage of basophils among CD33dim HLA DR- CD66b- cells (OR 1.03, 95% CI 1.004–1.055; p = 0.025), the percentage of naive CD4+ T cells among CD4+ T cells (OR 1.03, 95% CI 1.004–1.062; p = 0.025), the level of CD20 expression on IgD- CD38- B cells (OR 1.04, 95% CI 1.004–1.071, P = 0.027), the absolute count of CD66b++ myeloid cells (OR = 1.04, 95% CI 1.008–1.069; p = 0.013), the level of CX3CR1 expression on CD14- CD16+ monocytes (OR 1.05, 95% CI 1.012–1.085; p = 0.009), and the absolute count of NK cells (OR 1.05, 95% CI 1.003–1.096; p = 0.036) were identified as risk factors for VTE. The results described above are visualized in Figure 3. Following adjustment for multiple tests using the FDR method, none of the immune traits reached a significance level of 0.05.
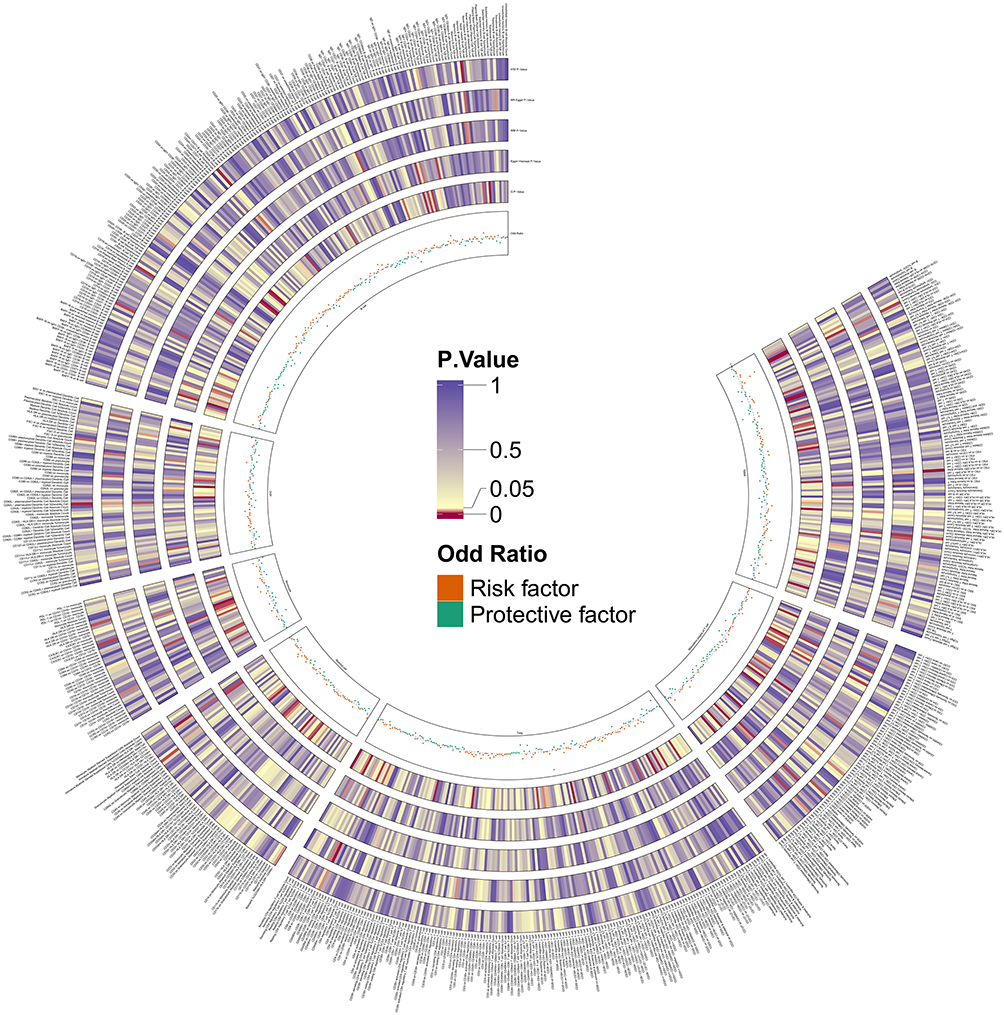 |
Figure 2 The circus plot presents the causal estimates and sensitivity analyses between Immune Cells and Venous Thromboembolism for the different MR methods. |
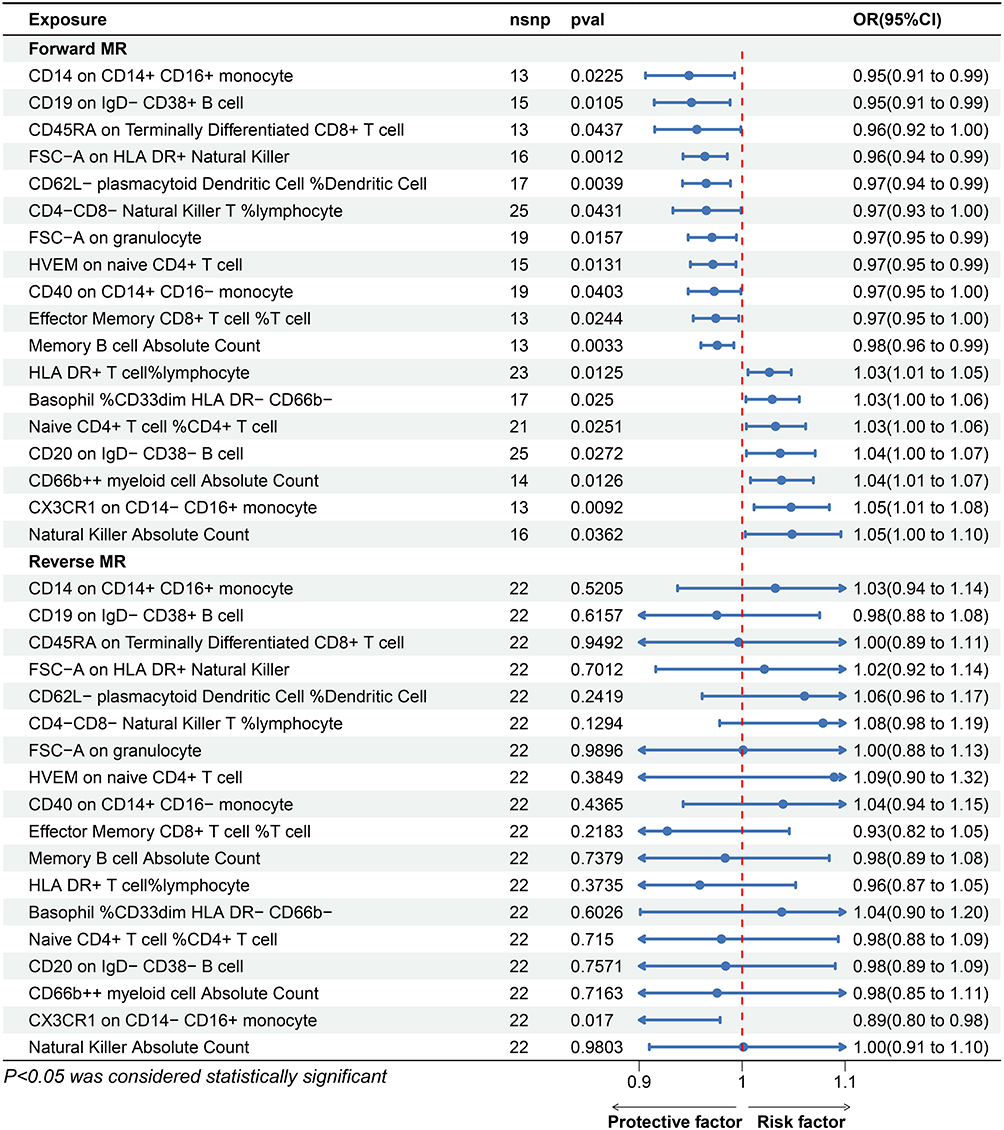 |
Figure 3 The bi-directional Mendelian randomization results between immunophenotypes and VTE for Protective factors. Abbreviations: MR, Mendelian randomization; OR, Odds ratio. |
Examination of Outliers and Heterogeneity
The results of scatter plots (Figures 4A–K and Figures 5A–G), funnel plots (Figures 6 and 7), and leave-one-out plots (Figures 8 and 9) did not show any outliers. Figure 10 presents the definitive results from the tests of heterogeneity, as well as for horizontal pleiotropy evaluation and Steiger test results. In the MR-PRESSO global test, the p values for all significant results were consistently > 0.05, indicating the absence of significant horizontal pleiotropy. The consistency in the findings was further confirmed through MR-Egger regression, aligning with the results obtained from the MR-PRESSO global test. Additionally, the outcomes of the Steiger test, featuring p-values significantly < 0.05, suggested a robust association between the IVs and immune phenotypes. These results confirmed the correct directionality of the causative effect, enhancing the reliability of MR insights into the causal linkage between immune cells and VTE.
 |
Figure 4 The scatter plots of the association between elven exposure and outcome with 5 methods (IVW approach, MR-Egger, simple mode, weighted mode, weighted median). The specific immune cell signature is shown above each subgraph (A-K). |
 |
Figure 5 The scatter plots of the association between seven exposure and outcome with 5 methods (IVW approach, MR-Egger, simple mode, weighted mode, weighted median). The specific immune cell signature is shown above each subgraph (A-G). |
 |
Figure 6 The funnel plots of the association between elven exposure and outcome. The specific immune cell signature is shown above each subgraph (A-K). |
 |
Figure 7 The funnel plots of the association between seven exposures and outcome. The specific immune cell signature is shown above each subgraph (A-G). |
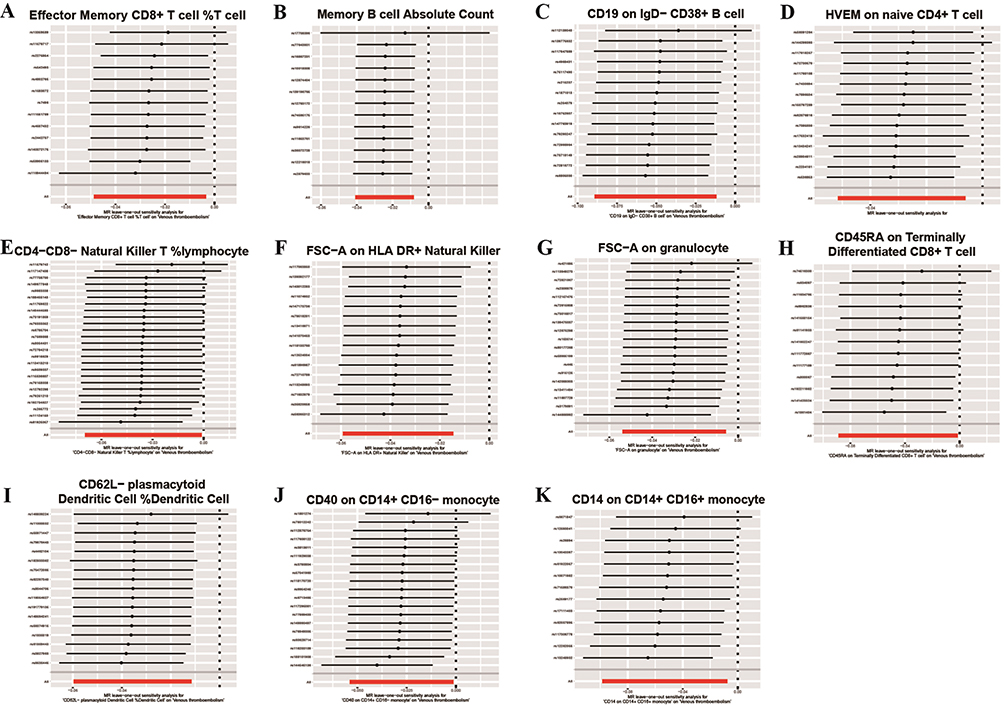 |
Figure 8 The leave-one-out analysis of the association between elven exposure and outcome. The specific immune cell signature is shown above each subgraph (A-K). |
 |
Figure 9 The leave-one-out analysis of the association between seven exposure and outcome. The specific immune cell signature is shown above each subgraph (A-G). |
 |
Figure 10 The results of sensitivity analyses. |
Reverse MR Analysis
We performed a reverse MR analysis by utilizing the IVW method to explore the causal relationship between the 18 immune phenotypes and VTE. Following the removal of LD, 23 SNPs were strongly linked with VTE, each demonstrating an F-statistic > 10.
The results indicate that 17 out of the 18 immunophenotypes did not demonstrate a statistically significant inverse relationship These included the level of CD14 expression in CD14+ CD16+ monocytes (p = 0.52), CD19 expression in IgD- CD38+ B cells (p = 0.62), CD45RA expression in terminally differentiated CD8+ T cells (p = 0.95), and FSC-A expression in HLA-DR + NK cells (p = 0.70). Similarly, the percentage of CD62L- plasmacytoid dendritic cells among dendritic cells (p = 0.24), the percentage of CD4-CD8- NK T cells among lymphocytes (p = 0.13), and FSC-A expression in granulocytes (p = 0.99) were not statistically significant. Additionally, HVEM expression on naive CD4+ T cells (p = 0.39), CD40 expression on CD14+ CD16- monocytes (p = 0.44), the percentage of effector memory CD8+ T cells among T cells (p = 0.22), and the absolute count of memory B cells (p = 0.53) did not exhibit statistically significant associations. Furthermore, the percentage of HLA DR+ T cells among lymphocytes (p = 0.37), the percentage of basophils among CD33dim HLA DR- CD66b- cells (p = 0.60), the percentage of naive CD4+ T cells among CD4+ T cells (p = 0.72), the expression levels of CD20 on IgD- CD38- B cells (p = 0.76), the absolute count of CD66b++ myeloid cells (p = 0.72), and the absolute count of NK cells (p = 0.98) did not show statistical significance. However, the expression level of CX3CR1 on CD14- CD16+ monocytes (p = 0.01) showed statistically significant associations, suggesting a potential bidirectional causal relationship. The reverse MR results are shown in Figure 2 and Supplementary Table 6.
Discussion
We conducted a comprehensive assessment of the causal associations between 731 immune cell traits and VTE through comparing large publicly available GWAS databases. To our knowledge, this is the first MR study to investigate the relationship between immune cells and VTE.
In this bidirectional two-sample MR analysis, we observed that increased percentages of naive CD4+ T cells among CD4+ T cells, CX3CR1 on CD14- CD16+ monocytes, HLA DR+ T cells among lymphocytes, and the absolute counts of NK cells, CD66b++ myeloid cells, along with the percentage of CD33dim HLA DR- CD66b- among basophils were associated with a heightened risk of VTE. Conversely, a higher genetically determined percentage of effector memory CD8+ T cells among T cells, HVEM on naive CD4+ T cells, CD45RA on terminally differentiated CD8+ T cells, CD40 on CD14+ CD16- monocytes, CD14 on CD14+ CD16+ monocytes, percentage of CD4-CD8- NK T cells among lymphocytes, FSC-A on HLA DR+ NK cells, the absolute count of memory B cells, CD19 on IgD- CD38+ B cells, FSC-A on granulocytes, and the percentage of CD62L- plasmacytoid dendritic cells among dendritic cells were associated with a reduced risk of VTE. Additionally, the reverse MR analysis indicated a significant association between CX3CR1 expression levels on CD14- CD16+ monocytes and VTE. These findings may lay the groundwork for future strategies in the prevention, diagnosis, and treatment of VTE.
The interaction between coagulation and inflammation has been known for several decades. Activation of the coagulation cascade and platelets can initiate immune responses including leukocyte recruitment, adhesion, extravasation, and activation. Simultaneously, the immune system influences the hemostatic system, a phenomenon known as immune thrombosis.28 Crosstalk between inflammation and hemostasis comprises intricate interactions involving proinflammatory cytokines, chemokines, adhesion molecules, tissue factor expression, and platelet and endothelial activation. Inflammatory processes upregulate procoagulant factors while concurrently inhibiting natural anticoagulant pathways and fibrinolytic activity, resulting in a prothrombotic tendency.29
T-cell activation is crucial for VTE, as it orchestrates the recruitment of neutrophils and monocytes. This process significantly impacts the formation and regression of thrombus neovascularization. Furthermore, CD4+ and CD8+ T cells have a functional role in facilitating the resolution of VTE.30
A study demonstrated an increased incidence of VTE in individuals infected with human immunodeficiency virus (HIV) and exhibiting reduced CD4+ T cell counts. This finding suggests a potential association between heightened hypercoagulability during progressive immunosuppression and the advancement of HIV disease.31 Our study discovered that the presence of HVEM on naive CD4+ T cells serves as a protective factor against VTE. HVEM, located on the surface of various cell types, both hematopoietic and non-hematopoietic, predominantly expresses in the spleen, thymus, bone marrow, lungs, and intestines.32 It has been reported that HVEM influences a range of immune responses, not only on mucosal surfaces but also in other parts of the body.33 However, the mechanisms underlying its association with thrombosis remain unclear.
Sustained interferon (IFN)-γ production during VTE requires the presence of NK cells.34 Some studies have suggested that HLA-DR+ NK cells exhibit a superior capacity for IFN-γ production upon cytokine stimulation when compared to their HLA-DR− counterparts.33 This finding indicates that NK cells expressing HLA-DR can promote VTE through IFN-γ production similar to T cells. HLA-DR-expressing NK cells play a role in frontline defense, engaging in crosstalk with the adaptive immune system through IFN-γ production, amplifying inflammation, and participating in cytotoxic responses, as well as the induction of T-cell activation. Our study found that FSC-A on HLA DR+ NK cells plays an important role in hinderi ng VTE or promoting VTE ablation. Currently, no clear experimental results are available to confirm whether NK cells influence VTE, and further studies are warranted.
The mechanism through which B cell affects VTE formation is unclear; however, Solveig et al suggested that genetically determined B cell depletion is a causative factor in VTE. In JHT mice, B cell deficiency is associated with systemic elevation of neutrophil and fibrinogen levels, indirectly contributing to thrombosis initiation.35
Monocytes regulate the immune response and perform multiple essential functions.36 They can be categorized into three subsets based on the expression of CD14 and CD16: classical monocytes (CD14+CD16-), intermediate monocytes (CD14+CD16+), and non-classical monocytes (CD14-CD16+).37 Different monocyte subsets play diverse roles in cardiovascular physiology and pathophysiology, although the underlying mechanisms are not well understood. Monocytes are the primary leukocytes involved in VTE resolution.38,39 A mouse experiment showed that Ly6CLo monocytes are not essential for thrombogenesis but are essential for normal VTE resolution;40 however, further research on the contribution of monocyte subsets is required.
Our study suggests that CX3CR1 on non-classical (CD14-CD16+) monocytes may increase the risk of VTE. CX3CR1 is classified as a seven-transmembrane G-protein coupled receptor which is expressed in vascular endothelial cells and monocytes/macrophages.41 The primary function of CX3CR1 in immune cells is to detect and infiltrate inflamed tissue, guided by the CX3CL1 gradient, thereby initiating an innate immune response.42 Thus, the interplay between CX3CL1 and CX3CR1 represents an initial phase in the host’s defense mechanism involving monocytes crawling or “patrolling” within the blood vessel lumen. In patients with Wegener’s granulomatosis, the expression of CX3CR1 in peripheral blood mononuclear cells has been suggested to enhance inflammation by facilitating the migration of leukocytes into inflammatory lesions.43 Simultaneously, our results also revealed their inverse relationship. This may be associated with the repeated stimulation and activation of CX3CR1 during inflammatory activation through the formation of neurovascular unit loops. Considering the complex and close relationship between the immune system and inflammation, this phenotype remains particularly noteworthy.
Plasmacytoid dendritic cells, a subset of dendritic cells, are highlighted for their significant role in innate immunity. Functioning as innate immune sentinels, plasmacytoid dendritic cells play essential roles in both infection immunity and autoimmunity. Their involvement in inflammation at specific sites includes the production of IFN-I and other mediators, such as CXCL4.44 In a study elucidating inflammation-induced thrombosis, findings indicated that platelets could incite inflammation by activating dendritic cells, thereby instigating VTE.26 Additionally, Maitre et al demonstrated that blood-isolated dendritic cells exhibited a preferential removal of platelets situated at the periphery of a preformed thrombosis.45 CD62L plays a pivotal role in regulating the recruitment of monocytes from the bloodstream to lymphoid tissues during inflammation.46 Our study suggests that the percentage of CD62L- plasmacytoid dendritic cells among dendritic cells is associated with a protective effect against VTE. However, additional evidence is warranted to substantiate this finding.
Previous studies have demonstrated that CD4+ T cells play a significant role in both thrombosis formation and resolution. One study observed the presence of CD4+ T cells at various stages of thrombus formation and resolution. Depletion of CD4+ T cells resulted in a reduction of macrophage numbers within the thrombus, decreased expression of the fibrinolytic marker urokinase-type plasminogen activator (uPA), and diminished activity of matrix metalloproteinase-9 (MMP-9). These findings suggest that CD4+ T cells have a functional role in the resolution phase of VTE.47,48
Additionally, CD39, an ectonucleotidase expressed on endothelial cells, plays a crucial role in inhibiting platelet function. Research indicates that CD39 hydrolyzes extracellular ATP and ADP, thereby reducing platelet activation and aggregation, which suppresses thrombus formation.49,50
Part of our findings align with former studies, but additional research is essential to identify the precise pathways that mediate these effects in humans.
It is important to note that the biological mechanisms underlying these associations are not fully understood. This study’s findings should therefore be interpreted with caution. While MR methods help mitigate confounding, the lack of a clear understanding of immune-mediated pathways in human thrombosis makes definitive causal conclusions challenging. Further research is needed to clarify the biological mechanisms linking immune cells to VTE.
Our study had some limitations. First, due to the absence of individual information in the original data, only summary statistics were evaluated, and a stratified analysis was not conducted. Second, only SNPs associated with known confounders were excluded; thus, other unknown confounders may have influenced our results. Finally, the GWAS data were derived from populations of European ethnicity, and caution should be exercised when generalizing our findings to other ethnicities.
Conclusion
Our study identified 18 types of immune cell signatures that could impact VTE development, offering novel insights for future mechanistic and clinical studies in this field. Further studies to prospectively validate our findings are needed.
Abbreviations
cDCs, conventional dendritic cells; CI, confidence interval; FDR, false discovery rate; GWAS, Genome-Wide Association Studies; HLA, human leukocyte antigen; HIV, human immunodeficiency virus; HVEM, herpes virus entry mediator; IFN, interferon; IVs, instrumental variables; IVW, inverse variance-weighted; LD, linkage disequilibrium; MR, Mendelian randomization; NK, natural killer; OR, odds ratio; SNPs, single nucleotide polymorphisms; VTE, venous thromboembolism.
Data Sharing Statement
All data generated or analyzed during this study are included in this published article [and its supplementary information files].
Ethics Statement
This study used publicly available genome-wide association study (GWAS) statistics. And it is exempt from approval based on item 1 and 2 of Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects dated February 18, 2023, China. So no new ethical approval was required.
Article 32: The use of human information data or biological samples to carry out life science and medical research involving human beings in the following circumstances may be exempted from ethical review, so as to reduce unnecessary burden on scientific researchers and promote the development of life science and medical research involving human beings, if it does not cause harm to human beings, does not involve sensitive personal information or commercial interests.
Item 1: Research using publicly available data obtained legally or data generated through observation without interfering with public behavior.
Item 2: Research using anonymized information data.
Acknowledgments
The authors are sincerely grateful to the participants and investigators for making the summary statistics data publicly available.
Funding
This work was supported by the Sichuan Medical Association [grant number 2019HR45].
Disclosure
The author(s) of this work have nothing to disclose.
References
1. Klemen ND, Feingold PL, Hashimoto B, et al. Mortality risk associated with venous thromboembolism: a systematic review and Bayesian meta-analysis. Lancet Haematol. 2020;7(8):e583–e593. doi:10.1016/S2352-3026(20)30211-8
2. Raskob GE, Angchaisuksiri P, Blanco AN, et al. Thrombosis: a major contributor to global disease burden. Arterioscler Thromb Vasc Biol. 2014;34(11):2363–2371. doi:10.1161/ATVBAHA.114.304488
3. Goldhaber SZ, Visani L, De Rosa M. Acute pulmonary embolism: clinical outcomes in the International Cooperative Pulmonary Embolism Registry (ICOPER). Lancet. 1999;353(9162):1386–1389. doi:10.1016/S0140-6736(98)07534-5
4. Kearon C. Natural history of venous thromboembolism. Circulation. 2003;107(23 Suppl 1):I22–I30. doi:10.1161/01.CIR.0000078464.82671.78
5. Rabinovich A, Kahn SR. The postthrombotic syndrome: current evidence and future challenges. J Thromb Haemost. 2017;15(2):230–241. doi:10.1111/jth.13569
6. Grosse SD, Nelson RE, Nyarko KA, Richardson LC, Raskob GE. The economic burden of incident venous thromboembolism in the United States: a review of estimated attributable healthcare costs. Thromb Res. 2016;137:3–10. doi:10.1016/j.thromres.2015.11.033
7. Barco S, Woersching AL, Spyropoulos AC, Piovella F, Mahan CE. European Union-28: an annualised cost-of-illness model for venous thromboembolism. Thromb Haemost. 2016;115(4):800–808. doi:10.1160/TH15-08-0670
8. Najem MY, Couturaud F, Lemarié CA. Cytokine and chemokine regulation of venous thromboembolism. J Thromb Haemost. 2020;18(5):1009–1019. PMID: 32020753. doi:10.1111/jth.14759
9. Vazquez-Garza E, Jerjes-Sanchez C, Navarrete A, Joya-Harrison J, Rodriguez D. Venous thromboembolism: thrombosis, inflammation, and immunothrombosis for clinicians. J Thromb Thrombolysis. 2017;44(3):377–385. PMID: 28730407. doi:10.1007/s11239-017-1528-7
10. Khorana AA, Mackman N, Falanga A, et al. Cancer-associated venous thromboembolism. Nat Rev Dis Primers. 2022;8(1):11. PMID: 35177631. doi:10.1038/s41572-022-00336-y
11. Emdin CA, Khera AV, Kathiresan S. Mendelian randomization. JAMA. 2017;318(19):1925–1926. doi:10.1001/jama.2017.17219
12. Burgess S, Scott RA, Timpson NJ, Davey Smith G, Thompson SG. Using published data in Mendelian randomization: a blueprint for efficient identification of causal risk factors. Eur J Epidemiol. 2015;30(7):543–552. doi:10.1007/s10654-015-0011-z
13. Davies N, Holmes M, Davey Smith G. Reading Mendelian randomisation studies: a guide, glossary, and checklist for clinicians. BMJ. 2018;362:k601. doi:10.1136/bmj.k601
14. Kurki M, Karjalainen J, Palta P, et al. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature. 2023;613(7944):508–518. doi:10.1038/s41586-022-05473-8
15. Orrù V, Steri M, Sidore C, et al. Complex genetic signatures in immune cells underlie autoimmunity and inform therapy. Nature Genet. 2020;52(10):1036–1045. doi:10.1038/s41588-020-0684-4
16. Sidore C, Busonero F, Maschio A, et al. Genome sequencing elucidates Sardinian genetic architecture and augments association analyses for lipid and blood inflammatory markers. Nature Genet. 2015;47(11):1272–1281. doi:10.1038/ng.3368
17. Sanna S, van Zuydam N, Mahajan A, et al. Causal relationships among the gut microbiome, short-chain fatty acids and metabolic diseases. Nature Genet. 2019;51(4):600–605. doi:10.1038/s41588-019-0350-x
18. Levin M, Judy R, Gill D, et al. Genetics of height and risk of atrial fibrillation: a Mendelian randomization study. PLoS Med. 2020;17(10):e1003288. doi:10.1371/journal.pmed.1003288
19. Dai H, Hou T, Wang Q, et al. Causal relationships between the gut microbiome, blood lipids, and heart failure: a Mendelian randomization analysis. Eur J Prev Cardiol. 2023;30(12):1274–1282. doi:10.1093/eurjpc/zwad171
20. Liu X, Qi X, Han R, Mao T, Tian Z. Gut microbiota causally affects cholelithiasis: a two-sample Mendelian randomization study. Front Cell Infect Microbiol. 2023;13:1253447. doi:10.3389/fcimb.2023.1253447
21. Kamat M, Blackshaw J, Young R, et al. PhenoScanner V2: an expanded tool for searching human genotype-phenotype associations. Bioinformatics. 2019;35(22):4851–4853. doi:10.1093/bioinformatics/btz469
22. Chen S, Yan S, Aiheti N, et al. A bi-directional Mendelian randomization study of sarcopenia-related traits and type 2 diabetes mellitus. Front Endocrinol. 2023;14:1109800. doi:10.3389/fendo.2023.1109800
23. Mao R, Yu Q, Li J. The causal relationship between gut microbiota and inflammatory dermatoses: a Mendelian randomization study. Front Immunol. 2023;14:1231848. doi:10.3389/fimmu.2023.1231848
24. Verbanck M, Chen C, Neale B, Do R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nature Genet. 2018;50(5):693–698. doi:10.1038/s41588-018-0099-7
25. Guo Z, Zhang T, Yun Z, et al. Assessing the causal relationships between human blood metabolites and the risk of NAFLD: a comprehensive Mendelian randomization study. Front Genetics. 2023;14:1108086. doi:10.3389/fgene.2023.1108086
26. Shang W, Qian H, Shen X, Wen Z, Zhang S, Chen D. Human blood metabolites and risk of severe COVID-19: a Mendelian randomization study. Eur J Clin Invest. 2023;53(11):e14079. doi:10.1111/eci.14079
27. Chen S, He W. Metabolome-wide Mendelian Randomization assessing the causal relationship between blood metabolites and bone mineral density. Calcif Tissue Int. 2023;112(5):543–562. doi:10.1007/s00223-023-01069-0
28. Han Z, Liu Q, Li H, et al. The role of monocytes in thrombotic diseases: a review. Front Cardiovasc Med. 2023;10:1113827. doi:10.3389/fcvm.2023.1113827
29. Aksu K, Donmez A, Keser G. Inflammation-induced thrombosis: mechanisms, disease associations and management. Curr Pharm Des. 2012;18(11):1478–1493. doi:10.2174/138161212799504731
30. Mukhopadhyay S, Gabre J, Chabasse C, Bromberg JS, Antalis TM, Sarkar R. Depletion of CD4 and CD8 positive T cells impairs venous thrombus resolution in mice. Int J Mol Sci. 2020;21(5):1650. doi:10.3390/ijms21051650
31. Tang Y-F, Wang Y, Xue T-J, et al. Clinical characteristics of HIV-infected patients with venous thromboembolism and different CD4+ T lymphocyte levels. J Inflamm Res. 2022;15:613–620. doi:10.2147/JIR.S346200
32. Doherty TA, Soroosh P, Khorram N, et al. The tumor necrosis factor family member LIGHT is a target for asthmatic airway remodeling. Nature Med. 2011;17(5):596–603. doi:10.1038/nm.2356
33. Erokhina SA, Streltsova MA, Kanevskiy LM, Telford WG, Sapozhnikov AM, Kovalenko EI. HLA-DR+ NK cells are mostly characterized by less mature phenotype and high functional activity. Immunol Cell Biol. 2018;96(2):212–228. doi:10.1111/imcb.1032
34. Cherian SM, Bobryshev YV, Inder SJ, Lord RS, Ashwell KW. Dendritic cells in venous pathologies. Angiology. 1999;50(5):393–402. doi:10.1177/000331979905000505
35. Hasselwander S, Xia N, Mimmler M, et al. B lymphocyte-deficiency in mice promotes venous thrombosis. Heliyon. 2022;8(11):e11740. doi:10.1016/j.heliyon.2022.e11740
36. Wolf AA, Yáñez A, Barman PK, Goodridge HS. The Ontogeny of Monocyte Subsets. Front Immunol. 2019;10:1642. doi:10.3389/fimmu.2019.01642
37. Ziegler-Heitbrock L, Ancuta P, Crowe S, et al. Nomenclature of monocytes and dendritic cells in blood. Blood. 2010;116(16):e74–e80. doi:10.1182/blood-2010-02-258558
38. Saha P, Humphries J, Modarai B, et al. Leukocytes and the natural history of deep vein thrombosis: current concepts and future directions. Arterioscler Thromb Vasc Biol. 2011;31(3):506–512. doi:10.1161/ATVBAHA.110.213405
39. Wakefield TW, Myers DD, Henke PK. Mechanisms of venous thrombosis and resolution. Arterioscler Thromb Vasc Biol. 2008;28(3):387–391. doi:10.1161/ATVBAHA.108.162289
40. Kimball AS, Obi AT, Luke CE, et al. Ly6CLo monocyte/macrophages are essential for thrombus resolution in a murine model of venous thrombosis. Thromb Haemost. 2020;120(2):289–299. doi:10.1055/s-0039-3400959
41. Kim K-W, Vallon-Eberhard A, Zigmond E, et al. In vivo structure/function and expression analysis of the CX3C chemokine fractalkine. Blood. 2011;118(22):e156–e167. doi:10.1182/blood-2011-04-348946
42. Hamon P, Loyher P-L, Baudesson de Chanville C, Licata F, Combadière C, Boissonnas A. CX3CR1-dependent endothelial margination modulates Ly6Chigh monocyte systemic deployment upon inflammation in mice. Blood. 2017;129(10):1296–1307. doi:10.1182/blood-2016-08-732164
43. Bjerkeli V, Damås JK, Fevang B, Holter JC, Aukrust P, Frøland SS. Increased expression of fractalkine (CX3CL1) and its receptor, CX3CR1, in Wegener’s granulomatosis—possible role in vascular inflammation. Rheumatology. 2007;46(9):1422–1427. doi:10.1093/rheumatology/kem168
44. Reizis B. Plasmacytoid dendritic cells: development, regulation, and function. Immunity. 2019;50(1):37–50. doi:10.1016/j.immuni.2018.12.027
45. Maître B, Mangin PH, Eckly A, et al. Immature myeloid dendritic cells capture and remove activated platelets from preformed aggregates. J Thromb Haemost. 2010;8(10):2262–2272. doi:10.1111/j.1538-7836.2010.03983.x
46. Xu H, Manivannan A, Crane I, Dawson R, Liversidge J. Critical but divergent roles for CD62L and CD44 in directing blood monocyte trafficking in vivo during inflammation. Blood. 2008;112(4):1166–1174. doi:10.1182/blood-2007-06-098327
47. Ley K, Laudanna C, Cybulsky MI, Nourshargh S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nat Rev Immunol. 2007;7(9):678–689. PMID: 17717539. doi:10.1038/nri2156
48. Williams MR, Azcutia V, Newton G, Alcaide P, Luscinskas FW. Emerging mechanisms of neutrophil recruitment across endothelium. Trends Immunol. 2011;32(10):461–469. PMID: 21839681; PMCID: PMC3185121. doi:10.1016/j.it.2011.06.009
49. Atkinson B, Dwyer K, Enjyoji K, Robson SC. Ecto-nucleotidases of the CD39/NTPDase family modulate platelet activation and thrombus formation: potential as therapeutic targets. Blood Cells mol Dis. 2006;36(2):217–222. PMID: 16476557. doi:10.1016/j.bcmd.2005.12.025
50. Marcus AJ, Broekman MJ, Drosopoulos JH, et al. The endothelial cell ecto-ADPase responsible for inhibition of platelet function is CD39. J Clin Invest. 1997;99(6):1351–1360. PMID: 9077545; PMCID: PMC507951. doi:10.1172/JCI119294
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
A Bidirectional Two-Sample Mendelian Randomization Study of Genetic Causality Between Vitamin D Levels and Pemphigus
Wang Y, Cheng S, Que H
Clinical, Cosmetic and Investigational Dermatology 2025, 18:1167-1176
Published Date: 10 May 2025
Causal Effects of Immune Cells on Reproductive Ill-Health, Including Abnormal Spermatozoa, Polycystic Ovary Syndrome and Spontaneous Abortion: Mendelian Randomization Analyses
Chen S, Sun S, Zhou Z, Zhou Z, Zhang R, Song W, Xin H, Yang Q, Dai S, Huang K, Niu W, Shi H, Guo Y
Journal of Multidisciplinary Healthcare 2025, 18:3219-3232
Published Date: 6 June 2025