")
Back to Journals » Journal of Inflammation Research » Volume 17
Combination of Methotrexate and Resveratrol Reduces Pro-Inflammatory Chemokines in Human THP-1 Cells
Authors Al-Nasser MM, Al-Saeedi MJ, Alhowaiti SA, Shinwari Z, Alhamlan FS, Alothaid H , Alkahtani S , Al-Qahtani AA
Received 2 August 2024
Accepted for publication 8 October 2024
Published 2 November 2024 Volume 2024:17 Pages 8085—8098
DOI https://doi.org/10.2147/JIR.S482503
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Subashchandrabose Chinnathambi
Moonerah M Al-Nasser,1 Mashael J Al-Saeedi,2 Saltana A Alhowaiti,2 Zakia Shinwari,3 Fatimah S Alhamlan,2,4 Hani Alothaid,5 Saad Alkahtani,1 Ahmed A Al-Qahtani2,4
1Department of Zoology, College of Science, King Saud University, Riyadh, Saudi Arabia; 2Department of Infection and Immunity, Research Centre, King Faisal Specialist Hospital & Research Centre, Riyadh, Saudi Arabia; 3Therapeutics & Biomarker Discovery for Clinical Applications, Stem Cell & Tissue Re-Engineering Program, King Faisal Specialist Hospital and Research Centre, Riyadh, Saudi Arabia; 4Department of Microbiology and Immunology, College of Medicine, Alfaisal University, Riyadh, Saudi Arabia; 5Department of Basic Sciences, Faculty of Applied Medical Sciences, Al-Baha University, Al-Baha, Saudi Arabia
Correspondence: Ahmed A Al-Qahtani, Email [email protected]
Introduction: Methotrexate (MTX) is a widely used anti-metabolite drug in cancer therapy, but its efficacy is often hindered by reactive oxygen species (ROS)-induced cellular toxicity. Resveratrol, a natural polyphenol, possesses antioxidant and anticancer properties. Therefore, this in vitro study aimed to investigate the synergistic anti-proliferative and anti-inflammatory effects of MTX and resveratrol in human THP-1 cells.
Methods: THP-1 cells were differentiated into macrophage-like cells using phorbol 12-myristate 13-acetate. In vitro experiments assessed the impact of various concentrations of MTX and resveratrol on cell viability and proliferation using the MTT assay. Concentration-effect curves were generated to elucidate their relationship. Gene expression was analyzed by RT-qPCR, while chemokine expression was measured via ELISA. Phagocytic and migratory activities were also evaluated.
Results: Differentiated THP-1 cells were treated with MTX and resveratrol and stimulated with dimethyl sulfoxide (DMSO) and lipopolysaccharide (LPS) as inflammatory stimuli. The combination of MTX and resveratrol exhibited an anti-proliferative effect in THP-1 cells (p = 0.03). The concentration-effect curve revealed IC50 values of 49.15 μg at 24 hours (R = 0.8236) and 2.029e-005 μg at 48 hours (R = 0.97) for MTX, and 20.17 μg at 48 hours (R = 1.000) and 55.38 μg at 96 hours (R = 0.9666) for resveratrol. Co-treatment with MTX and resveratrol significantly reduced mRNA and chemokine expression of CCL2, CCL3, CCL4, CCL5, and CXCL10 (p < 0.05). Moreover, decreased phagocytic and migratory activities were confirmed by phagocytosis and migration assays (p < 0.05).
Conclusion: The combination of MTX and resveratrol effectively attenuated pro-inflammatory activity in THP-1 cells, as evidenced by the downregulation of mRNA and chemokine expression. These findings suggest that the synergistic effects of MTX and resveratrol hold promise for enhancing cancer therapeutics.
Keywords: immunomodulatory, immune cells, anti-inflammatory, THP-1, methotrexate, resveratrol
Introduction
Methotrexate (MTX), an anti-inflammatory and anti-metabolic drug,1 is widely used as a therapeutic agent in a number of inflammatory diseases such as inflammatory bowel disease,2 psoriasis,3 rheumatoid arthritis,1 systemic lupus erythematosus,4 transplantation,5 and malignancies.6 The immunomodulatory effectiveness of MTX stems from its intricate and multifaceted mechanisms, which influence the immune system and induce changes. MTX possesses the ability to antagonize crucial enzymes involved in the biogenesis of purines and pyrimidines, thereby limiting the turnover of cancerous cells.7 MTX also mediates the release of cellular adenosine, which in turn enhances its anti-inflammatory activity by interacting with and inhibiting neutrophils, lymphocytes, and macrophages.8–10 Moreover, MTX stimulates the production of reactive oxygen species (ROS) by inhibiting dihydrofolate reductase, thereby depleting tetrahydrobiopterin, an essential cofactor of nitric oxide (NO).11 Additionally, it downregulates matrix metalloproteinases such as MMP-1, MMP-3, MMP-9, MMP-13, and TIMP-1.12 MTX also inhibits the nuclear factor-κB (NF-κB) signaling pathway which is responsible for inducing immune responses by promoting the production of IL-6, IL-1β, and MMPs.13 Furthermore, MTX halts the JAK/STAT signaling pathway, crucial for the upregulated expression of pro-inflammatory chemokines in inflammatory diseases.14
Macrophages are essential components of the immune system, strategically positioned within tissues to fulfill various biological functions.15 Most importantly, they orchestrate adaptive immune response by engulfing, neutralizing, and presenting foreign antigens.16 They generate pro-inflammatory and anti-inflammatory chemokines to recruit other immune cells to respond to infection and injury so as to prevent hyperresponsive immune system17 Nevertheless, macrophages have also been implicated in cellular events that lead to chronic inflammation, carcinogenesis, and tumor progression.18,19 For instance, a handful of studies have correlated inflammation in rheumatoid arthritis with elevated macrophage count.20–22
Altered macrophage differentiation has been demonstrated to affect tumour progression23,24 Despite the well-established anti-proliferative effects of MTX on tumor cells, its anti-inflammatory activity on macrophages requires further elucidation. A study by Municio et al (2018) conducted both in vitro and in vivo experiments and reported that MTX attenuated the proinflammatory responses of macrophages by upregulating the expression of A20, a suppressor of NF-κB. This resulted in a reduced inflammatory response of macrophages to pathogenic agents such as lipopolysaccharide (LPS) and synovial fluid from rheumatoid arthritis (RA) patients.25 In another recent study by Fuentelsaz-Romero et al (2021), investigating the anti-inflammatory properties of MTX, it was found that MTX suppresses the gene expression of human macrophages by inhibiting one carbon metabolism (OCM). This mechanism primes macrophages to become tolerant to lipopolysaccharide (LPS), a key trigger of the inflammatory process.26
Resveratrol (3,5,4’-trihydroxytransstilbene) is a polyphenolic compound found naturally in various edible sources such as mulberries, grapes, and peanuts. Over the past few decades, several in vitro and in vivo research studies have investigated the diverse biological properties of resveratrol. These include its anti-inflammatory, antioxidant, anti-atherogenic, anti-platelet, estrogen-like growth-enhancing, growth-antagonizing, immunomodulatory, and chemopreventive effects.27–29 Resveratrol is a strong anti-oxidant agent and possesses free radical scavenging activity owing to its property to stimulate diverse anti-oxidant enzymes.30
Regarding the synergistic effects of MTX and resveratrol on macrophages, no relevant studies have been conducted thus far. Only a limited number of studies have focused on elucidating the role of resveratrol in reversing MTX-induced cellular damage.27,28 A study conducted by Tunali-Akbay et al (2010) revealed elevated levels of serum TNF-α following administration of MTX. Subsequent investigation demonstrated that resveratrol was able to reduce these elevated TNF-α levels.27 Studies have shown that MTX promotes oxidative tissue injury by increasing the production of free radicals. This in turn triggers leukocyte accumulation and activates neutrophils, leading to the release of enzymes such as myeloperoxidase. These enzymes further exacerbate the secretion of free radicals, contributing to tissue damage.31
This in vitro study aimed to investigate the immunomodulatory functions of MTX alone and in combination with resveratrol, along with the associated molecular changes, using the THP-1 cell line. Specifically, the objectives were to: (1) assess the synergistic effects of MTX and resveratrol on monocyte cell proliferation, (2) determine the inhibitory activity (IC50) of MTX and resveratrol in THP-1 cells, (3) measure the mRNA expression levels of various proinflammatory chemokines in MTX- and resveratrol-treated THP-1 cells, (4) observe the effects of MTX and resveratrol on the expression of proinflammatory chemokines in THP-1 cells, (5) assess the effect of the combination of MTX and resveratrol on phagocytic activity, and (6) explore the effect of co-administration of MTX and resveratrol on migratory activity in macrophages. In this study, we treated THP-1 cells with MTX alone or in combination with resveratrol, employing various cellular assays to scrutinize biological changes.
Materials and Methods
Culture and Growth of THP-1 Cells
Human THP-1 monocytic cells (ATCC, Manassas, Virginia, USA) were cultivated in RPMI 1640 medium (Catalog number: 21875034, Thermo Fisher Scientific,Waltham, MA, USA) complemented with 10% complement-inactivated fetal bovine serum (FBS) (Catalog number: 10100147, Thermo Fisher Scientific,Waltham, MA, USA), 1% Pen Strep (10,000μg/mL streptomycin, 10,000 Units/mL Penicillin) (Catalog number: 15140122, Thermo Fisher Scientific, Waltham, MA, USA), and 1% of 200mM L-Glutamine (Catalog number: G7513, Sigma-Aldrich, St. Louis, MO, USA). The cells were then kept at a temperature of 37°C, 5% CO2, and 95% air.
THP-1 Differentiation into Macrophage-Like Cells
To differentiate THP-1 cells into macrophage-like cells, treatment with 100 ng/mL of phorbol 12-myristate 13-acetate for 48 hours in 6-well cell culture plates was performed (Catalog number: P1585,Sigma-Aldrich, St. Louis, MO, USA). Undifferentiated cells were then separated by rinsing with serum-free RPMI 1640 medium before conducting every experiment. For stimulation, THP-1 cells were exposed to 100 ng/mL of lipopolysaccharide (LPS) from Escherichia coli O26:B6, a gram-negative bacteria (Catalog number:L5543, Sigma-Aldrich, St. Louis, MO, USA) for 48 hours.
Treatment of Macrophage-Like Cells with Methotrexate and Resveratrol
Differentiated THP-1 cells (macrophage-like cells) were treated with dimethyl sulfoxide (DMSO) (Catalog number: D8418, Sigma-Aldrich, St. Louis, MO, USA) as a control for group 1 and 100 ng/mL of lipopolysaccharide (LPS) as a control for group 2. Furthermore, THP-1 cells were also treated with MTX (5µg/mL) alone or in combination with resveratrol (50 µg/mL and 100 µg/mL) and 100 ng/mL of LPS. Values denote the mean of triplicate experiments performed after 48 hours of incubation.
MTT Assay for Cell Proliferation
For cell viability and proliferation, the MTT assay was performed following the manufacturer’s protocol (Catalog number: G3582,Promega, Madison, Dane, USA). THP-1 cells were exposed to MTX (5µg/mL) alone or in combination with resveratrol (50µg/mL and 100µg/mL). Briefly, serum-containing media were replaced with serum-free media and MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] reagent. Cells were then placed into 96-multiwell cell culture plates at a volume of 5×104 cells per well and incubated at a temperature of 37°C for 48 hours. Next, the CellTiter 96® AQueous One Solution Reagent was added to every well and incubated for a further 4 hours at 37°C. Absorbance was detected at 490nm through a 96-well microtiter™ microplates reader (ThermoFisher Scientific™, USA). All experiments were performed as a set of three experiments. Cell proliferation was expressed as absorbance at 490nm with controls (vehicle untreated cells) for different time points.
Concentration-Effect Curve
The concentration-effect curve was constructed to elucidate the relationship between MTX and resveratrol and its corresponding effect, represented by the percentage of inhibition. The curve serves as a graphical representation of the drug’s potency and efficacy. The inhibitory concentration 50% (IC50) value, a critical parameter derived from this curve, signifies the concentration at which 50% inhibition of the intended biological response occurs. The IC50 value serves as a crucial metric for quantifying the potency of the drug and provides valuable insights into its pharmacological activity.
Expression Analysis
RT-qPCR analysis was performed to determine the gene expression. Total RNA was extracted from cultured THP-1 cells by utilizing the QIAamp RNA Blood Mini Kit (Catalog number: 52304,Qiagen, Hilden, Germany) according to the manufacturer’s guide. Followed by cDNA synthesis using All-in-One cDNA Synthesis SuperMix (Catalog number: 11766050, Thermo Fisher Scientific, Waltham, MA, USA). The generated cDNA was used for real-time PCR experiments. The mRNA levels were measured.32–36 Assays for gene expression used in this study were obtained from ThermoFisher Scientific (Catalog number: 4331182,Waltham, MA, USA) and included: CCL2 (Hs00234140_m1), CCL3 (Hs00234142_m1), CCL4 (Hs99999148_m1), CCL5 (Hs00982282_m1), and CX0CL10 (Hs00171042_m1). Afterwards, TaqMan Fast Advanced qPCR Master Mix (Catalog number: A28522, Applied Biosystems) was set up and amplification reaction was performed through Applied Biosystems 7900HT Fast Real Time PCR System. Relative gene expression data were analyzed using the ΔCt method. The mRNA expression of investigated genes was normalized to Beta-actin (ACTB). The mRNA expression data were shown as change in mean ± standard deviation (SD) relative to the control (DMSO and LPS treated) cells.
Enzyme-Linked Immunosorbent Assay (ELISA)
The ELISA assay was performed to determine the production of chemokine via Quantikine® ELISA Kit (Catalog number: DCP00, DMA00, DMB00, DRN00B and DY266,R&D Systems, Minneapolis, MN, USA) as per manufacturer’s guidelines. Each well of 96-well cell culture plates was supplemented with 100μL of cell culture supernatants and incubated for 2 hours at room temperature. Afterward, wells were rinsed four times with subsequent addition of primary antibodies and incubation for 2 hours at a room temperature. Then, HRP-Conjugated Streptavidin (100μL) was added to the wells and incubated for a further 20 minutes. This was followed by the addition of 100μL of the chromogenic substrate and further incubation in the dark for 30 minutes at room temperature. The process was concluded by the addition of stop solution (50μL) to every well. Absorbance was detected at optical density of 450 nm through a 96-well microtiter™ microplates reader (BioTek Instruments, Winooski, VT, USA). At last, the protein concentration was quantified through BioTek Gen5 Data Analysis Software.
Phagocytosis Assay
The phagocytic activity was observed through the Phagocytosis Assay Kit (Zymosan Substrate) (Catalog number: ab211156,Abcam, Cambridge, UK). MTX and resveratrol-treated THP-1 cells were incubated with Zymosan suspension for 75 minutes. The THP-1 cells were kept under the medium of serum-free RPMI at 5% CO2 and 37°C temperature. Afterward, the cells were rinsed thrice with a serum-free RPMI medium, followed by treatment of each well with 100μL of fixative to prevent external particles from degrading the specimen. Finally, absorbance was detected at an optical density of 405nm using the SpectraMax® MiniMax™300 Multi-Mode Detection Imaging Cytometer (Molecular Devices, CA, USA).
Migration Assay
Cell migration ability was assessed using the Transwell plates (Catalog number: ab235693, Abcam, Cambridge, UK). In total, 1×105 cells were placed on the Transwell chamber and serum-free culture medium (200 μL) was included. The lower partition of the chamber was supplied with 10% fetal calf serum and varied combinations and concentrations of MTX (5μg/mL) and resveratrol (50μg/mL and 100μg/mL). Next, after 48 hours, residual cells in the upper partition of the chamber were eliminated and the cells under the surface were stained with 0.5% crystal violet for 10 minutes and then fixation was done with 70% ethanol. Finally, a total of six fields of vision (×100 magnification) were decided at random and used to monitor cell migration.
Statistical Analysis
Research data were analyzed using GraphPad Prism 9.0 (GraphPad Software, Inc, La Jolla, CA, USA) and were presented as means ± standard deviation (SD) and standard error of the mean (SEM) using error bars. To check the level of significance and differences between untreated and treated samples, analysis of variance (ANOVA) was performed followed by a post hoc test (Bonferroni’s test). The P values will be set at *P<0.05, **P<0.005, ***P<0.001, and ****P<0.0001 to be considered statistically significant.
Results
Phagocytosis Study Using Light Microscopy
To assess phagocytic activity, heat-killed zymosan particles were introduced to the culture in serum-containing medium. Light microscopy analysis clearly depicted the engulfment of zymosan particles by the macrophage cells (Figure 1).
 |
Figure 1 Light microscopy of macrophage differentiated THP1 cells after exposure to Zymosan particles indicating that the particles were engulfed by the cells. |
Synergistic Effects of MTX and Resveratrol on Monocyte Cell Proliferation
To investigate the impact of MTX and resveratrol on monocyte cell proliferation, a MTT assay was conducted. Cells were treated with 5μg/mL of MTX alone or in combination with 50 or 100μg/mL of resveratrol. Analysis revealed that co-administration of 5μg/mL of MTX and 100μg/mL of resveratrol did not significantly alter monocyte cell proliferation compared to untreated control cells. However, both 5μg/mL of MTX alone (p = 0.0203) and in combination with 50μg/mL of resveratrol (p = 0.0493) significantly reduced monocyte cell proliferation at each time point (see Figure 2).
 |
Figure 2 Effects of the combination of methotrexate (MTX) and resveratrol on THP-1 cell proliferation. The graph demonstrates the time-dependent effect of 5 µg/mL MTX on cell proliferation after incubation with different concentrations of resveratrol (50 and 100 µg/mL) from 0 to 10 days after exposure. Each data point represents the mean of three independent replicates. Each point is the mean of triplicates for three independent experiments. Statistical significance was established using anova analysis with Bonferroni post-test. * is p= < 0.05. |
Determination of Inhibitory Activity (IC50) of MTX and Resveratrol in THP-1 Cells
The viability of THP-1 cells was assessed using the MTT assay, with treatments consisting of varying concentrations of MTX and resveratrol over 24, 48, and 96-hour periods. Concentration-effect curves were generated to determine % inhibition as a function of drug concentration, aiding in the calculation of IC50 values (the concentration required to induce 50% cell death). Figure 3A–D depicts the impact of MTX and resveratrol on THP-1 cells, revealing differing responses over time. The IC50 values for MTX were 49.15µg at 24 hours (R = 0.823) and 2.029e-005µg at 48 hours (R = 0.97), while those for resveratrol were 20.17 µg at 48 hours (R = 1.000) and 55.38 µg at 96 hours (R = 0.9666). These findings suggest that the response of THP-1 cells to MTX and resveratrol treatment varies over time. The observed differences in IC50 values between MTX and resveratrol at different time points may reflect distinct kinetics of drug action or varying sensitivities of THP-1 cells to each compound.
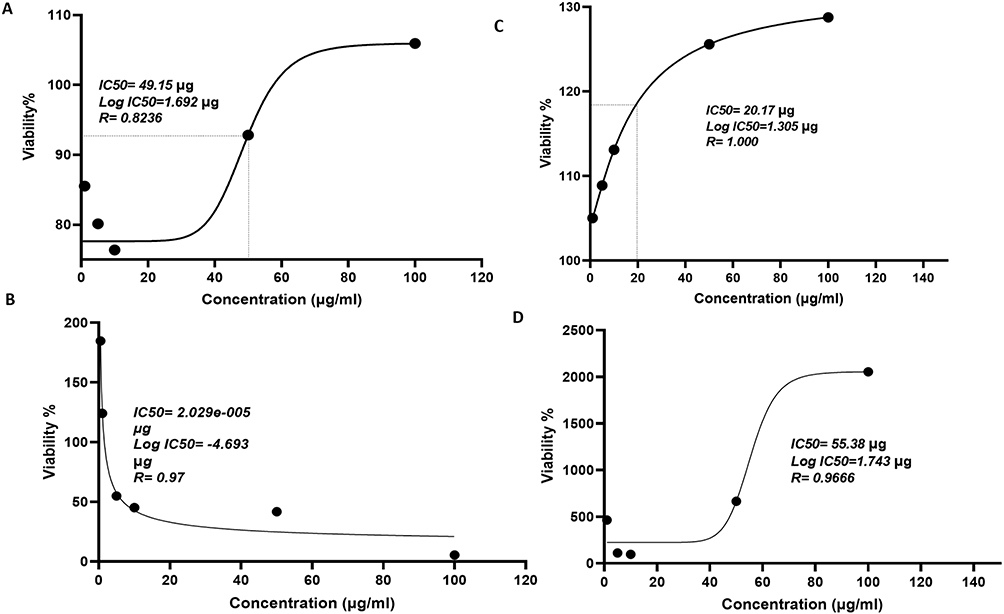 |
Figure 3 Concentration response curve for Methotrexate and Resveratrol in THP1-differentiated macrophages. The graph illustrates the concentration-effect relationship for Methotrexate (MTX) and Resveratrol, showing the percentage inhibition of cell growth as a function of drug concentration after 24 h exposure. The IC50 values represent the concentrations at which 50% inhibition of growth occurs. Panels (A and B) display the IC50 values for Methotrexate, while panels (C and D) present the IC50 values for Resveratrol. |
mRNA Expression Levels of Various Proinflammatory Chemokines in MTX and Resveratrol Treated THP-1 Cells
To assess the mRNA expression levels of proinflammatory chemokines (CCL2, CCL3, CCL4, CCL5, and CXCL10) in THP-1 cells treated with LPS and/or MTX alone or in combination with resveratrol, qRT-PCR analysis was employed. The results indicated an insignificant increase in the expression of CCL2 mRNA when cells were treated with 5 μg/mL of MTX and 50 or 100 µg/mL of resveratrol compared to control cells treated with DMSO (p = >0.9999 and p = 0.3667, respectively). Conversely, CCL2 mRNA expression significantly decreased when treated with 100 ng/mL LPS in combination with 5 μg/mL of MTX and 50 or 100 µg/mL of resveratrol compared to LPS-treated control cells (p = 0.0004 and p = 0.0007, respectively) (Figure 4a).
 |
Figure 4 mRNA expression levels of chemokines in THP-1 cells. The results depict the mRNA expression levels of chemokines in THP-1 cells post exposure to MTX, resveratrol, LPS, or a combination of the stimulants as shown in the graphs. (a) mRNA expression level of CCL2, (b) mRNA expression level of CCL3, (c) mRNA expression level of CCL4, (d) mRNA expression level of CCL5, and (e) mRNA expression level of CXCL10. The samples were processed in triplicate for three independent experiments, and the data are presented as mean values ± SEM. Statistical significance was established using Anova analysis with Bonferroni post-test. * is p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. |
Similarly, CCL3 mRNA expression increased significantly with 5 μg/mL of MTX and 100 µg/mL of resveratrol compared to DMSO control cells (p = 0.0002). However, it decreased significantly when treated with 100 ng/mL LPS in combination with 5 μg/mL of MTX and 50 or 100 µg/mL of resveratrol compared to LPS-treated control cells (p = 0.0147 and p = 0.0168, respectively) (Figure 4b). Furthermore, CCL4 mRNA expression substantially increased with 100 ng/mL LPS treatment compared to DMSO-treated control cells and with 5 μg/mL of MTX and 100 µg/mL of resveratrol when 5 μg/mL of MTX alone was considered as control (p > 0.9999). Conversely, it significantly decreased when treated with 100 ng/mL LPS in combination with 5 μg/mL of MTX and 50 or 100 µg/mL of resveratrol compared to LPS-treated control cells (p = 0.0027 and p = 0.0034, respectively) (Figure 4c).
Moreover, CCL5 mRNA expression significantly increased with 5μg/mL of MTX with or without 100 ng/mL of LPS compared to DMSO or LPS-treated control cells (p = 0.0005 and p < 0.0001, respectively). Conversely, it significantly decreased when treated with 100 ng/mL of LPS in combination with 5 μg/mL of MTX and 50 or 100 µg/mL of resveratrol compared to LPS-treated control cells (p = 0.1 and p = 0.0002, respectively) (Figure 4d).
Lastly, CXCL10 mRNA expression increased significantly with 5μg/mL of MTX compared to DMSO-treated control cells (p = 0.0022). However, it decreased with co-administration of 5 μg/mL of MTX and 50 or 100 µg/mL of resveratrol compared to MTX alone (p = 0.0076 and p = 0.0086, respectively). Similarly, it decreased upon treatment with 100 ng/mL of LPS along with 5 μg/mL of MTX and 50 or 100 µg/mL of resveratrol compared to LPS-treated control cells (p = 0.0005 and p = 0.0005, respectively) (Figure 4e).
Effects of MTX and Resveratrol on Expression of Proinflammatory Chemokines in THP-1 Cells
The ELISA assay was employed to quantify the expression of various chemokine proteins in THP-1 cells subjected to different treatments. The results demonstrated a significant increase in CCL2 expression upon treatment with 100 ng/mL of LPS compared to DMSO-treated control cells (p = 0.0009). However, treatment with 5µg/mL of MTX and 50 or 100 µg/mL of resveratrol, with or without LPS, led to a significant decrease in CCL2 expression compared to DMSO-treated control cells (p = 0.0022 and p = 0.0009, respectively) and LPS-treated control cells (p ≤ 0.0001) (Figure 5a). Similarly, CCL3 expression significantly decreased with 100 ng/mL of LPS treatment compared to DMSO-treated control cells (p = 0.0002). However, co-treatment with 5µg/mL of MTX and 50 or 100µg/mL of resveratrol, in the presence of LPS, led to an increase in CCL3 expression compared to LPS-treated control cells (p = 0.0137 and p = 0.0778, respectively) (Figure 5b).
 |
Figure 5 ELISA analysis of macrophage cell culture supernatant. ELISA results show the concentrations of analytes in the macrophage cell culture supernatant following stimulation with DMSO as a control for group 1 and LPS as a control for group 2. Group 1 includes MTX (5 µg/mL) only or with additional concentrations of resveratrol (50 µg/mL and 100 µg/mL), while group 2 comprises MTX (5 µg/mL) only or with additional concentrations of resveratrol (50 µg/mL and 100 µg/mL), all with LPS (100 ng/mL). (a) expression level of CCL2/MCP-1, (b) expression level of CCL3/MIP-1α, (c) expression level of CCL4/MCP-1, (d) expression level of CCL5, and (e) expression level of CXCL10. All samples were processed in triplicate for three independent experiments, and the data are presented as mean values ± SEM. Statistical significance was established using Anova analysis with Bonferroni post-test. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. |
Furthermore, treatment with 100 ng/mL of LPS significantly increased CCL4 expression compared to DMSO-treated control cells (p = 0.0002). However, all other concentrations led to a decrease in CCL4 expression compared to DMSO-treated control cells. Interestingly, co-treatment with 5 µg/mL of MTX and 50 or 100 µg/mL of resveratrol, in the presence of LPS, significantly increased CCL4 expression compared to the control group (p ≤ 0.0014) (Figure 5c).
Regarding CCL5 expression, treatment with 5µg/mL of MTX and 50 or 100 µg/mL of resveratrol significantly decreased expression compared to DMSO-treated control cells (p ≤ 0.0093), whereas co-treatment with 100 ng/mL of LPS and 5µg/mL of MTX significantly increased expression compared to LPS-treated control cells (p = 0.0023) (Figure 5d). Exposure to 100 ng/mL of LPS significantly increased CXCL10 expression compared to DMSO-treated control cells. However, co-administration of 100 ng/mL of LPS with 5µg/mL of MTX and 50 or 100 µg/mL of resveratrol significantly decreased expression compared to the group treated with 100 ng/mL of LPS and 5µg/mL of MTX alone (p < 0.0001) (Figure 5e).
Combination of MTX and Resveratrol Significantly Decreases Phagocytic Activity in Macrophages Compared to DMSO (Group 1) and LPS (Group 2) Controls
The results from the phagocytosis assay using Zymosan particles revealed that macrophage cells treated with 5μg/mL of MTX along with 50 or 100 μg/mL of resveratrol exhibited significantly reduced phagocytic activity compared to DMSO-exposed cells (control) (p = 0.0352 and p = 0.0458, respectively). Similarly, macrophage cells treated with 100 ng/mL of LPS also showed significantly decreased phagocytic activity compared to the control (p = 0.0325). Interestingly, the addition of 5 μg/mL of MTX to 100 ng/mL LPS-treated macrophage cells resulted in a significant increase in phagocytic activity (p = 0.0001). Moreover, although macrophage cells treated with LPS along with 5 μg/mL of MTX in combination with 50 or 100 μg/mL of resveratrol exhibited higher phagocytic activity than those treated with LPS alone (p = 0.0007 and p = 0.0019, respectively) (Figure 6).
 |
Figure 6 Phagocytosis assay using Zymosan particles. Macrophage cells were subjected to a phagocytosis assay using Zymosan particles. Group 1 was treated with DMSO as a control, comprising MTX (5 µg/mL) only or with additional concentrations of resveratrol (50 µg/mL and 100 µg/mL). Group 2, which serves as another control, was treated with LPS (100 ng/mL), including MTX (5 µg/mL) only or with additional concentrations of resveratrol (50 µg/mL and 100 µg/mL), all with LPS (100 ng/mL). The values represent the mean of experiments conducted in triplicate after 48 hours of incubation. Statistical significance was determined using Anova analysis with Bonferroni post-test. * is p < 0.05, **p < 0.01, ***p < 0.001. |
Co-Administration of MTX and Resveratrol Negatively Affects Migratory Activity in Macrophages Compared to DMSO (Group 1) and LPS (Group 2) Controls
The cell migration assay revealed significant differences between cells exposed to 100 ng/mL of LPS and untreated control cells (p = 0.0038). Furthermore, macrophage cells treated with 5μg/mL of MTX in combination with either 50 (p < 0.0001) or 100μg/mL (p = 0.0002) of resveratrol exhibited markedly reduced cell migration capability compared to cells treated with DMSO (control). Similarly, significant reductions in cell migration were observed when 100 ng/mL LPS-treated cells were exposed to 5μg/mL of MTX with 50 (p = 0.0035) or 100μg/mL (p = 0.0302) of resveratrol, as compared to the LPS alone group (Figure 7).
 |
Figure 7 Migratory potential of THP-1 cells. The migratory potential of THP-1 cells was assessed using a migration assay. Group 1 was treated with DMSO as a control, consisting of MTX (5 µg/mL) only or with additional concentrations of resveratrol (50 µg/mL and 100 µg/mL). Group 2, which serves as another control, was treated with LPS (100 ng/mL), including MTX (5 µg/mL) only or with additional concentrations of resveratrol (50 µg/mL and 100 µg/mL), all with LPS (100 ng/mL). Statistical significance was determined using Anova analysis with Bonferroni post-test. * is p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. |
Discussion
Chemotherapy serves as the primary treatment modality for numerous types of cancer.37 MTX stands out among various chemotherapeutic agents as a potent anti-metabolic drug, often administered alone or in combination with other anti-neoplastic medications. Its versatility extends to the treatment of various malignancies, including but not limited to head and neck, lung, breast, esophagogastric, gastric, and prostate cancers, as well as Hodgkin’s lymphoma, childhood acute lymphoblastic leukemia, and osteosarcoma.33 Previous in vivo study has demonstrated that MTX reduces secretion of pro-inflammatory chemokines, indicating its anti-inflammatory effect in chronic inflammatory diseases such as rheumatoid arthritis.34 However, the efficacy of MTX in treatment is hindered by its tendency to induce cellular toxicity through reactive oxygen species (ROS) production.32,35,36 Hence, studies have suggested combining MTX with antioxidant compounds to mitigate its ROS-induced toxicity.32,35 Resveratrol, a natural polyphenol compound, possesses both antioxidant and anticancer properties.29 This research aimed to uncover the synergistic anti-proliferative and anti-inflammatory effects of MTX and resveratrol in human THP-1 cells activated with DMSO and LPS.
Imbalances in homeostasis, triggered by internal or external stimuli, could initiate a cascade of cellular, metabolic, and molecular processes, thereby revealing the status of the body’s inflammatory response.38 It is important to note that persistently activated macrophages during inflammation have been linked in numerous chronic inflammatory conditions.39 Activated macrophages exhibit elevated pro-inflammatory signalling such as NF-κB signalling.40 Furthermore, activated macrophages orchestrate immune responses by recruiting other immune cells to the site of injury through chemoattractant proteins such as CCL2, CCL3, CCL4, CCL5, and CXCL10.41–45 The transcriptional transformation in activated macrophages also allows for increased survival, proliferation, phagocytosis, and migration activity.46
Overall, the current study demonstrated that co-treatment with MTX and resveratrol of DMSO and LPS-stimulated THP-1 cells inhibited proliferation, migration, and phagocytosis and influenced the expression of numerous pro-inflammatory proteins, as evidenced by decreased mRNA and protein expression of CCL2, CCL3, CCL4, CCL5, and CXCL10. To the best of our knowledge, this is the first in vitro study to evaluate the anti-inflammatory effects of MTX and resveratrol together in DMSO and LPS-stimulated human THP-1 cells.
Monocyte-differentiated macrophages play a crucial role in the immune system, contributing to both the initiation and resolution of inflammatory responses.47 While the function of cytokines in inducing macrophage polarization is well-established, the role of chemokines in this process remains to be fully elucidated. In the present study, we observed a reduction in the production and expression of pro-inflammatory chemokines, particularly CCL2, upon treatment with a combination of MTX and resveratrol. This suggests that the combination treatment contributed to the diminished classical activation of macrophages. One possible explanation for this phenomenon could be the disruption of miR-9-mediated downregulation of Dusp6 and ERK-mediated signal transduction, mechanisms known to support macrophage activation.48 The ERK signaling pathway is reportedly associated with proinflammatory macrophage polarization.49 Our findings are in line with that of a previous study confirming that resveratrol substantially downregulates the expression of CCL2.50 A considerable decrease in GM-CSF, a growth and differentiating factor for pro-inflammatory macrophages, with the administration of resveratrol has also been reported.50 In support of this, another previous study has shown that the binding of resveratrol to CCL2 inhibits its chemotactic effect on macrophages.51 Asides the effect of resveratrol on CCL2 at the protein level, Resveratrol has been discovered to influence the expression of numerous genes, encompassing those responsible for cytokines, caspases, matrix metalloproteinases, adhesion molecules, and growth factors.52 It is thus likely that resveratrol inhibition of CCL2 expression coupled to the binding of resveratrol to CCL2 protein inhibited differentiation of THP1 into macrophages or as observed in this study may inhibit activation of the THP1-differentiated macrophages. This inadvertently inhibits the production of pro-tumor cytokines like TNF-α or VEGF subsequently starving cancer cells in the tumor microenvironment (TME) (Figure 8). This coupled with MTX role in inhibiting folate synthesis may be an important synergistic mechanism to effectively treat specific cancers that are responsive to cytokine activities.
 |
Figure 8 CCL2, expressed by various cellular components including immune cells and cancer cells, within the TME, plays a crucial role. Its impact extends beyond simple chemotactic effects on CCR2-expressing monocytes/macrophages, also encompassing non-chemotactic effects. Resveratrol may prevent tumor growth through binding with CCL2 molecule. This interaction may block CCL2 interaction with its receptor CCR2, preventing differentiation of monocytes to tumor associate macrophages (TAM), resulting in no secretion of pro-tumor cytokines. |
The process of phagocytosis is an important determinant of the effective clearance of infection-causing microorganisms, foreign antigens, and neoplastic cells. Phagocytosis is equally important for preserving tissue homeostasis.53 Our results underscore a decreased phagocytic response of LPS-stimulated THP-1 cells with the co-administration of MTX and resveratrol in DMSO. The release of chemokines is commonly stimulated by pro-inflammatory cytokines. For example, TNF-α induces the release of CXCL10 through the activation of the PI3K/Akt and p38 MAPK signaling pathways, as well as the downstream NF-κB signaling pathway.54 In our study, we observed a decrease in mRNA and protein expression of CXCL10, which may be attributed to the disruption in TNF-α signaling. Previous research has demonstrated that TNF-α signaling disruption can also attenuate phagocytic function.55
In the tumor microenvironment, the infiltration of immune cells is crucial for cancer prognosis. Chemokines play a pivotal role in mediating the migration of immune cells and supporting the survival and proliferation of cancer cells, as discussed earlier. These molecular mechanisms have significant implications for cancer progression and treatment outcomes.56,57 Interestingly, our findings revealed that co-administration of MTX and resveratrol effectively suppressed the migration of stimulated THP-1 cells. Notably, CCL2, CCL3, CCL4, CCL5, and CXCL10 are well-established chemoattractant proteins involved in cell migration.41–45 These chemokines play pivotal roles in recruiting immune cells to sites of inflammation or injury, as well as in promoting the migration of cancer cells within the tumor microenvironment.58 The ability of MTX and resveratrol to attenuate the migration of THP-1 cells suggests a potential therapeutic strategy for modulating immune cell trafficking and their role in inhibiting tumor metastasis.
In human THP-1 cells, the expression of CCL2 and CXCL10 was markedly suppressed upon co-treatment with MTX and resveratrol, providing evidence for the potential mechanism underlying the observed reduction in cell migration ability. Our results align with previous research indicating that treatment with resveratrol leads to a significant downregulation of CCL2 and CXCL10 expression.50 Notably, our study represents the first investigation into these underlying cellular responses in differentiated THP-1 cells following co-treatment with MTX and resveratrol. This novel exploration sheds light on the intricate interplay between these compounds and their effects on chemokine expression, offering valuable insights into potential therapeutic strategies for modulating immune cell behavior and inhibiting tumor metastasis.
Before concluding, here are few things worth discussing. Firstly, in this study, we treated THP-1 cells with MTX alone or in combination with resveratrol, employing various cellular assays to scrutinize biological changes. THP-1, an acute leukemia monocytic cell line derived from humans, is extensively utilized for investigating macrophage functions, signaling pathways, and drug-induced inflammatory and immune responses. Serving as a reliable in vitro model, THP-1 cells allow assessment of macrophage proliferation, differentiation, migration, and immune modulation in response to external stimuli. Recent studies have underscored the utility of THP-1 cell lines in elucidating immune-modulatory responses induced by natural compounds and conventional drugs, a notion corroborated by the findings of our current investigation. Secondly, the reason to investigate the effects of MTX and resveratrol at different time points was to understand the cellular responses to treatment over time. Through assessment of MTX at diverse time points, we planned to recognize the ideal “window for action” and gain insight of the temporality of its anti-inflammatory potential. Furthermore, we observed significant variation in IC50 values for MTX across different time points. This could be due to the kinetics of the MTX uptake by THP-1 cells, time-contingent response of cells, or likely metabolism in culture medium over time. Finally, we also witnessed that MTX and resveratrol combination did not significantly alter monocyte proliferation at higher concentration but they did at lower concentration. The plausible explanation of this could be dose-dependent effects of resveratrol. Higher doses of resveratrol with its antioxidant properties might have counteracted the anti-proliferative effects of MTX.
Conclusion
The results of our study indicate that the combination of MTX and resveratrol effectively attenuates pro-inflammatory activity in THP-1 cells, as evidenced by decreased mRNA and protein expressions of chemokines like CCL2, CCL3 and CCL5, as well as inhibited phagocytic and migratory activity of THP-1-differentiated macrophages. These findings suggest that the combined effects of MTX and resveratrol hold promise for enhancing cancer therapeutics through modulation of immune cell contribution to cancer cell trafficking and metastasis. However, we recognize the need for further molecular investigations to elucidate the mechanistic pathways underlying the synergistic inflammatory properties of MTX and resveratrol reported in our study. Such investigations will provide valuable insights into the potential mechanisms of action and optimize the therapeutic use of this combination in the treatment of inflammatory and neoplastic diseases.
Abbreviations
ACTB, Beta-actin; DMSO, dimethyl sulfoxide; ELISA, Enzyme-linked Immunosorbent Assay; FBS, fetal bovine serum; GM-CSF, Granulocyte-macrophage colony-stimulating factor; LPS, lipopolysaccharide; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; MTX, Methotrexate; NF-κB, nuclear factor-κB; NO, nitric oxide; OCM, one carbon metabolism; PMA, phorbol 12-myristate 13-acetate; RA, rheumatoid arthritis; ROS, reactive oxygen species; RT-qPCR, reverse transcription-quantitative polymerase chain reaction; SD, standard deviation; TME, tumor microenvironment; VEGF, vascular endothelial growth factor.
Acknowledgments
The authors extend their appreciation to the King Salman center For Disability Research for funding this work through Research Group no KSRG-2023-305.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Zhao Z, Hua Z, Luo X, et al. Application and pharmacological mechanism of methotrexate in rheumatoid arthritis. Biomed Pharmacother. 2022;150:113074. doi:10.1016/J.BIOPHA.2022.113074
2. Herfarth HH, Kappelman MD, Long MD, Isaacs KL. Use of methotrexate in the treatment of inflammatory bowel diseases. Inflamm Bowel Dis. 2016;22(1):224–233. doi:10.1097/MIB.0000000000000589
3. da Silva CAP, Von Kossel K, Leszczynski M, Melnik T, Riera R. Methotrexate for psoriasis. Cochrane Database Syst Rev. 2019;2019(4). doi:10.1002/14651858.CD010498.PUB2
4. McKeon KP, Jiang SH. Treatment of systemic lupus erythematosus. Aust Prescr. 2020;43(3):85–90. doi:10.18773/austprescr.2020.022
5. Xu Z, Mo X, Kong Y, et al. Mini-dose methotrexate combined with methylprednisolone as a first-line treatment for acute graft-versus-host disease: a phase 2 trial. J Transl Int Med. 2023;11(3):255–264. doi:10.2478/JTIM-2023-0111/HTML
6. Yang V, Gouveia MJ, Santos J, et al. Breast cancer: insights in disease and influence of drug methotrexate. RSC Med Chem. 2020;11(6):646–664. doi:10.1039/d0md00051e
7. Chan ES, Cronstein BN. Methotrexate–how does it really work? Nat Rev Rheumatol. 2010;6(3):175–178. PMID: 20197777. doi:10.1038/nrrheum.2010.5
8. Riksen NP, Barrera P, Van Den Broek PHH, Van Riel PLCM, Smits P, Rongen GA. Methotrexate modulates the kinetics of adenosine in humans in vivo. Ann Rheum Dis. 2006;65(4):465–470. doi:10.1136/ARD.2005.048637
9. Cronstein BN, Eberle MA, Gruber HE, Levin RI. Methotrexate inhibits neutrophil function by stimulating adenosine release from connective tissue cells. Proc Natl Acad Sci. 1991;88(6):2441–2445. doi:10.1073/PNAS.88.6.2441
10. Haskó G, Cronstein B. Regulation of inflammation by adenosine. Front Immunol. 2013;4(APR). doi:10.3389/FIMMU.2013.00085/FULL
11. Chalupsky K, Cai H. Endothelial dihydrofolate reductase: critical for nitric oxide bioavailability and role in angiotensin II uncoupling of endothelial nitric oxide synthase. Proc Natl Acad Sci U S A. 2005;102(25):9056–9061. doi:10.1073/PNAS.0409594102
12. Fiedorczyk M, Klimiuk PA, Sierakowski S, Gindzienska-Sieskiewicz E, Chwiecko J. Serum matrix metalloproteinases and tissue inhibitors of metalloproteinases in patients with early rheumatoid arthritis. J Rheumatol. 2006;33(8):1523–1529.
13. Tak PP, Firestein GS. NF-κB: a key role in inflammatory diseases. J Clin Invest. 2001;107(1):7–11. doi:10.1172/JCI11830
14. Hu X, li J, Fu M, Zhao X, Wang W. The JAK/STAT signaling pathway: from bench to clinic. Signal Transduc Target Therap. 2021;6(1):1–33. doi:10.1038/s41392-021-00791-1
15. Lee CZW, Ginhoux F. Biology of resident tissue macrophages. Development. 2022;149(8). doi:10.1242/DEV.200270/275252
16. Rosales C, Uribe-Querol E. Phagocytosis: a fundamental process in immunity. Biomed Res Int. 2017;2017:9042851. doi:10.1155/2017/9042851
17. Hirayama D, Iida T, Nakase H. The phagocytic function of macrophage-enforcing innate immunity and tissue homeostasis. Int J Mol Sci. 2018;19:92. doi:10.3390/IJMS19010092
18. Bingle L, Brown NJ, Lewis CE. The role of tumour-associated macrophages in tumour progression: implications for new anticancer therapies. J Pathol. 2002;196(3):254–265. doi:10.1002/PATH.1027
19. Steidl C, Lee T, Shah SP, et al. Tumor-associated macrophages and survival in classic Hodgkin’s lymphoma. N Engl J Med. 2010;362(10):875–885. doi:10.1056/NEJMoa0905680
20. Mulherin D, Fitzgerald O, Bresnihan B. Synovial tissue macrophage populations and articular damage in rheumatoid arthritis. Arthritis Rheum. 1996;39(1):115–124. doi:10.1002/ART.1780390116
21. Brennan FM, McInnes IB. Evidence that cytokines play a role in rheumatoid arthritis. J Clin Invest. 2008;118(11):3537–3545. doi:10.1172/JCI36389
22. Hamilton JA, Tak PP. The dynamics of macrophage lineage populations in inflammatory and autoimmune diseases. Arthritis Rheum. 2009;60(5):1210–1221. doi:10.1002/ART.24505
23. Lin EY, Nguyen AV, Russell RG, Pollard JW. Colony-stimulating factor 1 promotes progression of mammary tumors to malignancy. J Exp Med. 2001;193(6):727–740. doi:10.1084/jem.193.6.727
24. Hamilton JA. Colony-stimulating factors in inflammation and autoimmunity. Nat Rev Immunol. 2008;8(7):533–544. doi:10.1038/nri2356
25. Municio C, Dominguez-Soto Á, Fuentelsaz-Romero S, et al. Methotrexate limits inflammation through an A20-dependent cross-tolerance mechanism. Ann Rheum Dis. 2018;77(5):752–759. doi:10.1136/ANNRHEUMDIS-2017-212537
26. Fuentelsaz-Romero S, Barrio-Alonso C, García Campos R, et al. The macrophage reprogramming ability of antifolates reveals soluble CD14 as a potential biomarker for methotrexate response in rheumatoid arthritis. Front Immunol. 2021:12. DOI:10.3389/FIMMU.2021.776879/FULL
27. Tunali-Akbay T, Sehirli O, Ercan F, Sener G. Resveratrol protects against methotrexate-induced hepatic injury in rats. J Pharm Pharm Sci. 2010;13(2):303–310. doi:10.18433/j30k5q
28. El-Sheikh AAK, Morsy MA, Al-Taher AY. Protective mechanisms of resveratrol against methotrexate-induced renal damage may involve BCRP/ABCG2. Fundam Clin Pharmacol. 2016;30(5):406–418. doi:10.1111/FCP.12205
29. Salehi B, Mishra AP, Nigam M, et al. Resveratrol: a double-edged sword in health benefits. Biomedicines. 2018;6(3):91. doi:10.3390/biomedicines6030091
30. Malhotra A, Bath S, Elbarbry F. An organ system approach to explore the antioxidative, anti-inflammatory, and cytoprotective actions of resveratrol. Oxid Med Cell Longev. 2015;2015:1–15. doi:10.1155/2015/803971
31. Sullivan GW, Sarembock IJ, Linden J. The role of inflammation in vascular diseases. J Leukoc Biol. 2000;67(5):591–602. doi:10.1002/jlb.67.5.591
32. Soylu Karapinar O, Pinar N, Özcan O, Özgür T, Dolapçıoğlu K. Protective effect of alpha-lipoic acid in methotrexate-induced ovarian oxidative injury and decreased ovarian reserve in rats. Gynecological Endocrinol. 2017;33(8):653–659. doi:10.1080/09513590.2017.1306847
33. Koźmiński P, Halik PK, Chesori R, Gniazdowska E. Overview of dual-acting drug methotrexate in different neurological diseases, autoimmune pathologies and cancers. Int J Mol Sci. 2020;21(10):3483. doi:10.3390/ijms21103483
34. Barsig J, Yam G, Lehner MD, Beume R. Methotrexate treatment suppresses local cytokine and chemokine production in rat adjuvant arthritis. Drugs Exp Clin Res. 2005;31(1):7–11.
35. Kabakcı B, Kurutaş E, Bakariş S, Güngör M. The effect of erythropoietin on methotrexate-induced renal damage in rats: biochemical and histopathological studies. Cukurova Med J. 2015;40(1):98–106. doi:10.17826/cutf.23047
36. Wang W, Zhou H, Liu L. Side effects of methotrexate therapy for rheumatoid arthritis: a systematic review. Eur J Med Chem. 2018;158:502–516. doi:10.1016/j.ejmech.2018.09.027
37. Tilsed CM, Fisher SA, Nowak AK, Lake RA, Lesterhuis WJ. Cancer chemotherapy: insights into cellular and tumor microenvironmental mechanisms of action. Front Oncol. 2022;12. doi:10.3389/FONC.2022.960317/FULL
38. Chovatiya R, Medzhitov R. Stress, inflammation, and defense of homeostasis. Mol Cell. 2014;54(2):281–288. doi:10.1016/j.molcel.2014.03.030
39. Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol. 2008;8(12):958–969. doi:10.1038/nri2448
40. Dorrington MG, Fraser IDC. NF-κB signaling in macrophages: dynamics, crosstalk, and signal integration. Front Immunol. 2019;10(APR). doi:10.3389/FIMMU.2019.00705/FULL
41. AbuSamra DB, Panjwani N, Argüeso P. Induction of CXCL10-mediated cell migration by different types of galectins. Cells. 2021;10(2):274. doi:10.3390/cells10020274
42. Bhavsar I, Miller CS, Al-Sabbagh M. Macrophage inflammatory protein-1 alpha (MIP-1 alpha)/CCL3: as a biomarker. Gen Meth Biomark Res Their Applicat. 2015;1:223–249. doi:10.1007/978-94-007-7696-8_27
43. Murooka TT, Rahbar R, Platanias LC, Fish EN. CCL5-mediated T-cell chemotaxis involves the initiation of mRNA translation through mTOR/4E-BP1. Blood. 2008;111(10):4892–4901. doi:10.1182/blood-2007-11-125039
44. Sindhu S, Kochumon S, Shenouda S, Wilson A, Al-Mulla F, Ahmad R. The cooperative induction of CCL4 in human monocytic cells by TNF-α and palmitate requires MyD88 and Involves MAPK/NF-κB signaling pathways. Int J Mol Sci. 2019;20(18):4658. doi:10.3390/ijms20184658
45. Yoshimura T, Li C, Wang Y, Matsukawa A. The chemokine monocyte chemoattractant protein-1/CCL2 is a promoter of breast cancer metastasis. Cell Mol Immunol. 2023;20(7):714–738. doi:10.1038/s41423-023-01013-0
46. Bergman NH, Passalacqua KD, Gaspard R, Shetron-Rama LM, Quackenbush J, Hanna PC. Murine macrophage transcriptional responses to Bacillus anthracis infection and intoxication. Infect Immun. 2005;73(2):1069–1080. doi:10.1128/IAI.73.2.1069-1080.2005
47. Gordon S, Taylor PR. Monocyte and macrophage heterogeneity. Nat Rev Immunol. 2005;5(12):953–964. doi:10.1038/nri1733
48. Carson WF, Salter-Green SE, Scola MM, Joshi A, Gallagher KA, Kunkel SL. Enhancement of macrophage inflammatory responses by CCL2 is correlated with increased miR-9 expression and downregulation of the ERK1/2 phosphatase Dusp6. Cell Immunol. 2017;314:63–72. doi:10.1016/J.CELLIMM.2017.02.005
49. Través PG, de Atauri P, Marín S, et al. Relevance of the MEK/ERK signaling pathway in the metabolism of activated macrophages: a metabolomic approach. J Immunol. 2012;188(3):1402–1410. doi:10.4049/jimmunol.1101781
50. Schwager J, Richard N, Widmer F, Raederstorff D. Resveratrol distinctively modulates the inflammatory profiles of immune and endothelial cells. BMC Complement Altern Med. 2017;17(1):309. doi:10.1186/s12906-017-1823-z
51. Tripathi DK, Nagar N, Naidu G, et al. Mechanistic insights into inhibition of CCL2-guided leukocyte recruitment by oxyresveratrol and its derivatives. J Mol Liq. 2024:398. DOI:10.1016/J.MOLLIQ.2024.124218
52. Ghafouri-Fard S, Bahroudi Z, Shoorei H, et al. Disease-associated regulation of gene expression by resveratrol: special focus on the PI3K/AKT signaling pathway. Cancer Cell Int. 2022;22(1):298. doi:10.1186/s12935-022-02719-3
53. Martinez-Marin D, Jarvis C, Nelius T, Filleur S. Assessment of phagocytic activity in live macrophages-tumor cells co-cultures by confocal and Nomarski microscopy. Biol Methods Protoc. 2017;2(1). doi:10.1093/BIOMETHODS/BPX002
54. Wang Z, Ao X, Shen Z, et al. TNF-α augments CXCL10/CXCR3 axis activity to induce epithelial-mesenchymal transition in colon cancer cell. Int J Biol Sci. 2021;17(11):2683–2702. doi:10.7150/ijbs.61350
55. Sharma L, Wu W, Dholakiya SL, et al. Assessment of phagocytic activity of cultured macrophages using fluorescence microscopy and flow cytometry. Methods Mol Biol. 2014;1172:137–145. doi:10.1007/978-1-4939-0928-5_12
56. Chow MT, Luster AD. Chemokines in cancer. Cancer Immunol Res. 2014;2(12):1125–1131. doi:10.1158/2326-6066.CIR-14-0160
57. Burugu S, Asleh-Aburaya K, Nielsen TO. Immune infiltrates in the breast cancer microenvironment: detection, characterization and clinical implication. Breast Cancer. 2017;24(1):3–15. doi:10.1007/s12282-016-0698-z
58. Qin R, Ren W, Ya G, et al. Role of chemokines in the crosstalk between tumor and tumor-associated macrophages. Clin Exp Med. 2023;23(5):1359–1373. doi:10.1007/s10238-022-00888-z
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Blood Coral Polysaccharide Helps Prevent D-Gal/LPS-Induced Acute Liver Failure in Mice
Li C, Lai S, Yi R, Zhou X, Zhao X, Li Q
Journal of Inflammation Research 2022, 15:4499-4513
Published Date: 8 August 2022
Deciphering Natural Killer Cell Cytotoxicity Against Medulloblastoma in vitro and in vivo: Implications for Immunotherapy

Gauthier M, Pierson J, Moulin D, Mouginot M, Bourguignon V, Rhalloussi W, Vincourt JB, Dumas D, Bensoussan D, Chastagner P, Boura C, Decot V
ImmunoTargets and Therapy 2024, 13:319-333
Published Date: 26 June 2024