")
Back to Journals » Journal of Inflammation Research » Volume 18
Deciphering the Role of Innate Lymphoid Cells Group 3 in the Gut Microenvironment: A Narrative Review of Their Novel Contributions to Autoimmune Disease Pathogenesis
Authors Wang H, Wang T, He Z, Wen C, Huang L, Wang M
Received 18 December 2024
Accepted for publication 9 April 2025
Published 28 April 2025 Volume 2025:18 Pages 5741—5757
DOI https://doi.org/10.2147/JIR.S512652
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Xiaoyu Liu
Hongli Wang,1,2 Tengyue Wang,1,2 Zhixing He,1,2 Chengping Wen,1,2 Lin Huang,1,2 Mingzhu Wang1,2
1Research Institute of Chinese Medical Clinical Foundation and Immunology, School of Basic Medical Science, Zhejiang Chinese Medical University, Hangzhou, 310053, People’s Republic of China; 2Key Laboratory of Chinese Medicine Rheumatology of Zhejiang Province, Zhejiang Chinese Medical University, Hangzhou, 310053, People’s Republic of China
Correspondence: Lin Huang; Mingzhu Wang, Email [email protected]; [email protected]
Abstract: Type 3 Innate lymphoid cells (ILC3s) play a crucial role in intestinal immune function by serving as an innate effector that contributes to early-life defense against pathogens and helps protect the intestines from bacterial infections. ILC3s exert their immune function through cytokine secretion, patrolling actions and the generation of memory ILC3s that aid in repairing epithelial tissue and preserving mucosal barrier integrity. Moreover, dysregulation of ILC3s function has been implicated in the pathogenesis and progression of autoimmune diseases. This comprehensive review aims to explore the interactions between gut microbes, gut microbial metabolites, and diet in relation to ILC3s within the context of the gut microenvironment. Furthermore, the gut microenvironment has the potential to influence distant extra-intestinal sites through immunomodulation, thereby modifying their risk of inflammation. The gut has emerged as a significant focus of autoimmune disease research in recent years. However, the relationship between gut ILC3s and autoimmune diseases remains poorly understood. This paper aims to examine the potential association between ILC3s and autoimmune diseases.
Keywords: ILC3s, IL-22, short-chain fatty acids, rheumatoid arthritis, systemic lupus erythematosus, ankylosing spondylitis
Introduction
Innate Lymphoid cells (ILCs) represent a group of lymphocytes with important effector and regulatory functions in innate immunity and tissue remodeling, of which ILC3s is an important class.1 ILC3s are capable of producing IL-17, IL-22 and granulocyte-macrophage colony-stimulating factor (GM-CSF), and are recognized by their expression of the transcription factor RORγt.2 In addition, transcription factors can also clarify the identity of ILC3 and maintain its function, such as GATA3,3 Zbtb1,4 HIF-1,5 etc. ILC3s are predominantly localized on mucosal surfaces, including the gut, lung, and skin,6,7 serving as crucial innate effect in early-life defence against pathogens. ILC3s primarily produce IL-22 to safeguard intestinal immunity. IL-22 acts as a potent inducer of mucins, large glycoproteins that form a thick, normally impenetrable barrier on the mucosal surface.8 Additionally, IL-22 promotes the differentiation and proliferation of Paneth cells through the PI3K/AKT/mTOR axis.9 These cells secrete various antimicrobial molecules, including defensins and antimicrobial peptides (AMPs). ILC3s localization exhibit rapid responsiveness to environmental and pathogenic signals. Villous ILC3s, which are predominantly immobile during homeostasis, can acquire migratory “patrolling” properties and enhance cytokine expression in response to inflammation.10 During bacterial infections, ILC3s-derived IL-22 clears bacteria by regulating the expression of antimicrobial genes in the epithelium.11 Recent evidence suggests that ILC3s possess memory properties, and exposure to pathogenic bacteria results in the generation of a highly functional and persistent ILC3s subpopulation, akin to memory T cells. These memory-like ILC3s exhibit a preferential expansion upon re-encounter with pathogens and produce substantial amounts of IL-22, thereby contributing to long-term mucosal defense.12
ILC3s show a strong correlation with the pathogenesis of autoimmune diseases, such as inflammatory bowel disease (IBD).13 However, their mechanism of action in other autoimmune diseases such as Rheumatoid arthritis(RA),14 Systemic lupus erythematosus (SLE),15 are fully controversial. This article provides a thorough overview of the role of ILC3s and associated cytokines in maintaining intestinal immune homeostasis, the impact of the intestinal microenvironment on ILC3s functions that may contribute to the pathogenesis of autoimmune disease.
Cell Classification
Lymphocytes are categorized into two groups based on their ability to recognize specific antigens: acquired immune cells and ILCs.16 ILCs represent a group of lymphocytes with important effector and regulatory functions in innate immunity and tissue remodeling.1 This innate lymphocyte family includes various subpopulations, including Group 1 ILCs (ILC1s), Group 2 ILCs (ILC2s), Group 3 ILCs (ILC3s), lymphoid tissue inducers (LTi) and Natural killer (NK) cells.17 Each subtype of ILCs possesses distinct transcription factor requirements and cytokine production capacities, influencing their roles in various immune responses.18
ILC1s and NK cells constitute closely related subsets within the ILC family, distinguished by their expression of the transcription factor T-bet and their ability to produce Interferonγ (IFN-γ).19 Although NK cells and ILC1s have historically been considered as functionally distinct cell types, ILCs are primarily found residing within tissue, in contrast to NK cells, which are mainly circulating in the bloodstream.20,21 ILC2s secrete type 2 cytokines, such as IL-5 and IL-13, in response to cytokines like IL-33. These cytokines contribute to tissue repair and defense against helminth infections.22 ILC3s are recognized by their expression of the transcription factor RORγt and cytokines such as IL-17, IL-22, and granulocyte-macrophage colony-stimulating factor (GM-CSF).2 LTi cells, which emerge early in embryogenesis are involved in secondary lymphoid organogenesis, T-cell tolerance, and T- and B-cell function.23 Although it has been suggested that LTi could be categorized within ILC3s due to their mutual requirement for RORγt expression for both their development and function, LTi/LTi-like cells are now acknowledged as a distinct population. This distinction is based on their unique gene expression pattern and their development from precursors that are different from the precursors of all other ILCs and conventional natural killer (cNK) cells.24,25
ILC3s are derived from fetal liver and bone marrow progenitors that colonize the intestine during embryonic development and early postnatal life.24 ILC3s can be further classified into distinct subgroups with significant differences in tissue localization, transcriptional regulation, cell surface phenotypes and biological functions.The identification of subpopulations of ILC3s for mice is mainly based on the expression of two cellular markers, CD196 (CCR6) and CD335 (NKp46), and there are three basic populations of mouse ILC3s:NKp46+CCR6- (NCR+), NKp46-CCR6- (NCR-) and NKp46-CCR6+ (CCR6+ LTi-like) ILC3s.2,26 NCR+ ILC3s express mainly IL-22 and less IL-17, NCR-ILC3 expresses mainly IL-17 and less IL-22.27 Furthermore, NCR+ ILC3s exhibit transcriptional expression of T-bet, which is essential for the expression of NKp46 and the production of IFN-γby ILC1s.28 CCR6+ LTi-like cells are predominantly located within crypt plaques (CP) and mature isolated lymphoid follicles (ILF), and have been demonstrated to promote the formation of lymph nodes and Peyer’s patches.29
Transcription Factors
Transcription Growth Factors
ILC3s represent the predominant subpopulation of gut-resident ILCs. The development and function of ILC3s are dependent on transcription factors, including GATA3, RORgt, Zbtb1, Id2, and others3,30–32 (Table 1).
 |
Table 1 Transcription Factors Have Been Demonstrated to Promote the Development and Function of ILC3s |
GATA3
GATA3 is essential for the promotion of the development of multiple ILC subpopulations, including ILC3s, and for the maintenance of ILC1s and ILC2s.3 GATA3 plays a crucial role in maintaining the equilibrium of ILC3s through its influence on IL-7Rα expression, which is a pivotal process in the development, maintenance and proliferation of ILC3s. GATA3 binds to a region within intron 2 of the Il7r gene, exhibiting a binding pattern that is highly similar to that observed in ILC3s, ILC2s and Th2 cells.33 While GATA3 has been demonstrated to positively regulate the expression of genes specific to the NKp46+ ILC3s subset, it has also been shown to exert a negative regulatory effect on the expression of genes specific to the CCR6+ ILC3s subset and lymphoid tissue inducer (LTi)-specific genes in NKp46+ ILC3s. Furthermore, GATA3 is essential for the production of IL-22 in both LTi and NKp46+ ILC3s.34 In the absence of Gata3, there was a reduction in the expression of the antimicrobial peptide Regiiγby intestinal epithelial cells.33
Zbtb1
Zinc finger and BTB domain containing 1 (Zbtb1) plays a crucial role in lymphocyte development.4 Zbtb1 has been demonstrated to prevent DNA damage in cell lines by initiating trans-damage DNA synthesis in response to replication stress. Conversely, deletion of Zbtb1 has been observed to result in increased DNA damage and activation of p53-dependent apoptosis in immune progenitor cells, thereby affecting lymphocyte.40 Although Zbtb1 is commonly expressed in different ILC subpopulations, its presence has been shown to significantly impact the production of NKp46+ILC3s. Zbtb1 has been identified as a crucial transcription factor essential for the generation of NKp46+ ILC3s. The deletion of Zbtb1 has been observed to impair the production of IFN-γby ILC3s, while its absence does not affect the ILC3s’ capacity to secrete IL22.31
Nfil3
Nfil3 (also known as E4BP4) is a basic leucine zipper transcription factor that has been demonstrated to regulate a range of cellular processes in lymphocyte subpopulations. Nfil3 plays a pivotal role in the development of ILCs subpopulations, which are crucial for intestinal immunoprotection, as well as the high levels of Nfil3 expression observed in all ILC populations.37 Nfil3 is responsible for the development and/or maintenance of ILC3s in a cell-intrinsic manner. The expression of Nfil3 increases continuously from the CLP to the ILCP to the mature ILC3s stages, with the highest levels of Nfil3 observed in intestinal ILC3s. Nfil3 is indispensable for the progression from the CLP to ILCP stages. It can therefore be postulated that the expression of Nfil3 may precede that of RORγt and Gata3, and that it may regulate the expression of RORγt and Gata3.37,38
Maintenance Function
ILC3s represent the predominant subpopulation of gut-resident ILCs and are instrumental in the pathogenesis of infection and the process of tissue repair. However, the mechanisms by which they adapt to the intestinal environment to maintain tissue residency remain unclear, and some common transcription factors are summarized for reference in this section.41
HIF-1
One of the principal mechanisms of cellular adaptation to hypoxia is the activation of hypoxia-inducible factor (HIF)-1, which is expressed in the majority of cells within the immune system, as well as in the IEC.5 ILC3s are highly prevalent in the intestinal mucosa, and thus the elevation in RORγt expression and transcriptional capacity by HIF-1 facilitates the proliferation and activation of ILC3s under hypoxic conditions, which is crucial for maintaining barrier homeostasis.42 It has been demonstrated that hyperbaric oxygen therapy reduces the abundance of ILC3s and its IL-22 production, and stimulates the HIF-1α-IL-22 signalling axis in ILC3s, thereby impairing host immunity.43 Furthermore, HIF-1αplays a role in the conversion of ILC3s to ILC1s in NKp46+ cells. It has been demonstrated that HIF-1αcan maintain ILC3s status in intestinal NKp46+ cells by reducing T-bet expression when HIF-1αis absent.44 Genetic deletion of HIF-1αselectively within ILC3s progenitors results in impaired IL-17 and IL-22 secretion in vivo.40 This deficiency corresponds with reduced expression of epithelial target genes and heightened susceptibility to enteric microbial infections.40 In contrast, HIF-1α ablation solely within NCR+ILC3s precludes conversion to inflammatory ILC1s, thereby preventing dextran sodium sulfate colitis-exacerbated intestinal damage.38
C-Maf
C-Maf has been identified as a pleiotropic regulator of effector programming in adaptive and innate-like lymphocytes.45 A transcriptomic analysis of ILC subpopulations revealed that c-Maf was the most highly expressed gene in ILC3s,46 suggesting that c-Maf may have a function in this spectrum.
NCR+ ILC3s possess plasticity, undergoing full conversion to ILC1s status upon acquiring a type 1 effector profile.47 C-Maf establishes physiological equilibrium between type 1 and type 3 effector states in steady-state NCR+ ILC3s by suppressing type 1 signatures and limiting conversion to physiological ILC1s.47 Recent investigations employing ILCs network analyses have demonstrated that c-Maf plays a regulatory role in maintaining the equilibrium between ILC3s and ILC1s populations within the intestinal mucosa.48 C-Maf engages in a negative feedback loop with T-bet to solidify ILC3s identity, while also restricting functions of Runx3 and TCF-1 that may drive type 1 transition in wild-type NCR+ ILC3s.49
The NF-κB and Notch signalling pathways, which are activated by IL-1ß and IL-18, respectively, represent potent exogenous enhancers of c-Maf expression in CCR6-ILC3s. And C-Maf forms a negative feedback loop with T-bet to preserve the identity of CCR6 ILC3s.49 C-Maf has been demonstrated to enhance ILC3 accessibility and to bolster RORγt activity.
Gut Microenvironment and ILC3s
Establishing a balanced gut microbiota interacting homeostatically with the intestinal epithelial barrier proves integral to preserving normal enteric functioning.50 However, disturbances stemming from genetic, environmental, dietary shifts or inflammatory states can incite dysbiosis of the resident microbiota.51 Such disruptions impact crucial programs governing intestinal immunity, including ILC3s development and functional capacities.51
ILC3s subsets receive regulatory cues from dietary elements through discrete molecular circuits. The AhR, G protein-coupled receptors (GPCRs), and Retinoic acid receptors (RARs) transduce signals influencing ILC3s activation status and cytokine secretion profiles.52,53 AhR engagement mediates ILC3s responses to microbial tryptophan catabolites while GPCRs and RARs coordinate detection of microbial metabolites and retinoids, respectively.53
AhR
AhR functions as a key immune cellular sensor of environmental conditions through its dual roles as a cytoplasmic receptor and transcription factor.54 Within ILC3s subsets, AhR exhibits high expression and proves indispensable for development and homeostasis.54 AhR primarily binds endogenous ligands including tryptophan metabolites derived from gut microbiota, such as indole and indole-3-acetic acid.55,56 Such ligand engagement enhances ILC3s secretion of IL-22 while preventing intestinal epithelial cell transformation into a DNA damage response phenotype.56 AhR ligands may also exert direct cytoprotective effects on epithelial cells.57
The WASH protein complex exhibits elevated nuclear expression specific to ILC3s, where it proves necessary to maintain the subset independently of developmental requirements.58 Within these cells, WASH recruits chromatin remodeler Arid1a to the aryl hydrocarbon receptor promoter to activate gene transcription.58 Meanwhile, the IL-22 receptor exhibits wide distribution across intestinal stem cell membranes and transduces signals crucial for IL-22-mediated responses.59 L-fucose significantly increases IL-22R cryptal expression and may engage AhR pathways to promote intestinal IL-22 release59 (Figure 1).
 |
Figure 1 AhR is highly expressed in ILC3s and is required for ILC3s development and maintenance. The gut microbiota-derived tryptophan metabolites indole and indole-3-acetic acid bind to AhR to activate ILC3s and regulate IL-22 production. AhR ligand binds to AhR and enhances ILC3s to secrete IL-22 and prevent intestinal epithelial cell transformation through the DNA damage response (DDR). In ILC3s, WASH recruits Arid1a to the AhR promoter, thereby activating AhR expression. |
Ffar2
Activation of GPCRs, such as free fatty acid receptor 2 (Ffar2) and free fatty acid receptor3 (Ffar3), typically induces expansion of the peripheral ILCs population. Among colonic ILC3s subsets, Ffar2 defines a dominant signalling pathway through regulation of proliferation and function, acting primarily on NCR+ ILC3s.60 While dispensable for lymphoid tissue development, Ffar2 expression on ILC3s may contribute to increased abundance within colonic lymphoid aggregates.60 Ffar2 activates AKT or ERK signalling through the AKT and STAT3 axis and increases ILC3s-derived IL-22. In addition, the Ffar2 receptor stimulates the activation of PI3K, Stat3, Stat5 and mTOR, promoting colonic ILC3s activation.61 Activation of the Ffar2 receptor promotes colonic ILC3s proliferation in situ and, in the absence of Ffar2, reduces ILC3s proliferation in situ and the production of derived IL-22, leading to impaired intestinal epithelial function, alterations in mucus-associated proteins and antimicrobial peptides, and increased susceptibility to colonic injury and bacterial infection.62
Retinoic Acid Receptor
Dendritic cells act as the principal source of Retinoic acid receptors (RARs) in the intestine and metabolize vitamin A into its biologically active form, retinoic acid.63 Retinoic acid and vitamin A signaling induces a homing receptor switch allowing ILC3s and ILC1s migration to the intestine independent of retinoic acid effects on ILC2s homing.64 Within the intestinal compartment, retinoic acid functions synergistically with IL-2 and other stimuli to induce expression of the homing integrin α4β7 and IFN-γ production in ILC1s and ILC3s.65 Specifically, retinoic acid upregulates ILC3s abundance and IL-22 expression via promoting IL-1β and IL-22 generation in IL-3 exposed to IL-23.65,66 The cytokine response elicited demonstrates dose-dependency on retinoic acid levels.66 Collectively, these mechanisms fine-tune ILC3s developmental trajectories and functional capacities (Figure 2).
 |
Figure 2 Retinoic acid regulates ILC3s homing and promotes ILC3s growth. Retinoic acid induces a homing receptor switch that allows ILC3s and ILC1s to migrate to the intestine, but ILC2s migration to the intestine is not regulated by retinoic acid. Retinoic acid inhibits ILC2s but increases ILC3s numbers and IL-22 expression, mainly by promoting IL-22 production in IL-1β- and IL-23-stimulated ILC3s and by controlling LTi upstream of the transcription factor RORγt. |
SCFAs
Short-chain fatty acids (SCFAs) regulate ILC3s function by activating specific GPCRs, inhibiting histone deacetylases, stimulating histone acetyltransferases, stabilizing hypoxia-inducible factor 1, and GPR43-dependent metabolic regulation.67 Acetate and butyrate prove indispensable for colonic ILC3s proliferation and effector cytokine generation, respectively.52 Acetate activates Ffar2 on ILC3s and epithelia to boost IL-1β production, promoting neutrophil-ILC3s cross-talk. The acetate-Ffar2 axis elevates ILC3s IL-1R expression, heightening IL-1β sensitivity and inducing IL-22/IFN-γ secretion.61,67 Acetate inhibits ILC3s-derived IL-17A independently of Ffar2 through direct histone deacetylase inhibition as well.67 Under homeostasis, butyrate negatively regulates terminal ileal Peyer’s patch ILC3s frequency, correlating with enhanced antigen-specific mucosal immunity.68 Acetate promotes ILC3s proliferation primarily via Ffar2 and downstream AKT, STAT3 and mTOR signaling, whereas propionate preferentially induces IL-22 through Ffar369 (Figure 3).
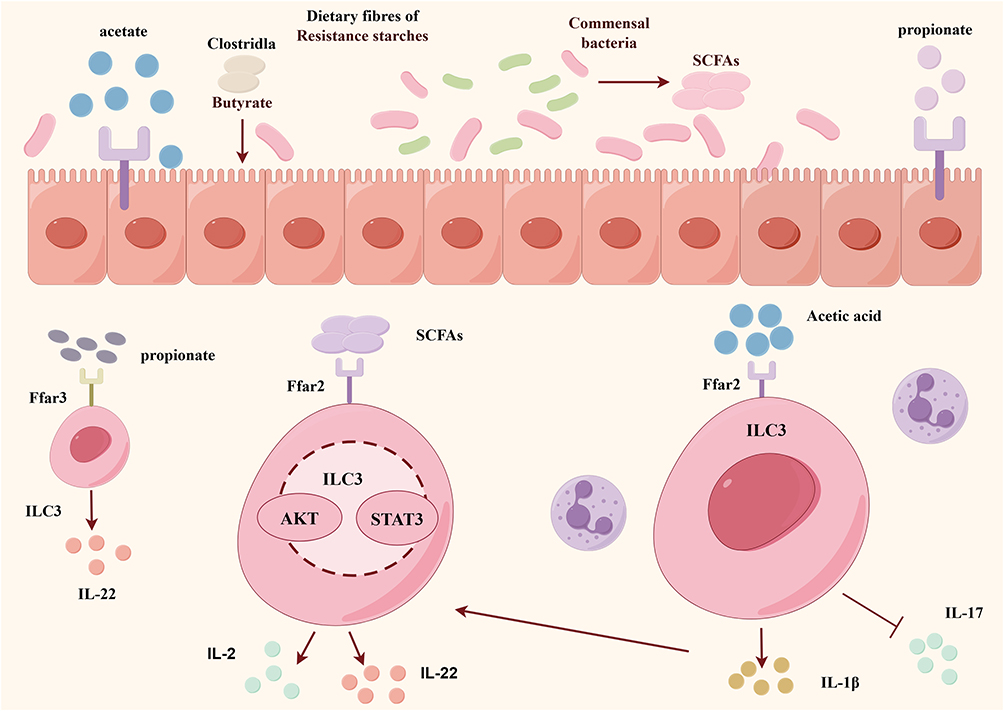 |
Figure 3 SCFAs are essential for the proliferation and effector function (IL-22 production) of colonic ILC3s to promote intestinal immunity. Gut microbiota ferment dietary fibre to produce SCFAs, which stimulate IL-22 production by ILC3s, mainly through activation of the AKT and STAT3 pathways. SCFAs, including acetate and butyrate, etc. Acetate activates FFAR2 to increase the production of IL-1β, which in turn increases ILC3s susceptibility to IL-1β, promotes IL-22 production, and also reduces ILC3s-induced IL-17A levels. |
Intestinal Micronutrients
Intestinal micronutrients such as vitamins A and D critically govern ILC3s maturation and functional preservation. Deficient adult vitamin A consumption hampers ILC3s proliferation and secondary lymphoid structure maturation.70 Specifically, reduced frequencies of intestinal ILC3s subsets exhibit diminished IL-22 production.70 Vitamin A proves essential to maintain intestinal ILC3s pools through ensuring cellular stability.57
Vitamin D arises from sunlight or diet and influences food allergy risk.71 The biologically active 1.25(OH)2D3 form engages the vitamin D receptor (VDR), highly expressed gut-resident ILC3s, to control lineage commitment and functionality.72 VDR deficiency or signaling blockade impairs ILC3s maturation and immunity particularly involving lymphoid tissue inducer cells.72 VDR governs IL-22 synthesis by regulating ILC3s proliferation and potentially IL-22 gene transcription independently of symbiotic bacteria.73 Vitamin D ensures colonic ILC3s-derived IL-22 and protective Th17 amplification during infection, as its deprivation elicits prolonged, severe enteric pathogenesis.73 While vitamin D clearly impacts ILC3s physiology, deeper exploration of underlying mechanisms remains necessary.
Gut Microorganisms
SFB
Segmented filamentous bacteria (SFB) are a group of host-adapted symbiotic organisms that are widely distributed in vertebrates, including humans and mice.74 SFB predominantly colonizes the terminal ileum in mice, where they typically instigate autoimmunity. SFB colonization induces transcriptomic shifts within the lamina propria, characterized by surging serum amyloid A 1 and 2 (SAA1/2) production in overlying epithelial cells.75 SFB engages IL-22-mediated STAT3 signaling in epithelial cells to induce SAA1/2 expression, implicating a homeostatic mechanism coupling barrier integrity maintenance to microbial colonization.76 As the principal intestinal source of IL-22, ILC3s express IL-22 at levels exceeding Th17 cells 10-fold.77 SFB also elicits REG3γ production and IL-22-dependent phosphorylation of STAT3 in epithelial cells, supporting epithelial renewal.78,79 IL-22 stimulated by SFB further induces futute-2 and fucosylation pathways enhancing paneth cell antimicrobial capacity.79
Dietary composition intricately shapes SFB behavior through regulatory interactions with ILC3s. Nutrient intake sustains SFB-associated epithelial morphology via IL-22 signaling, whereas IL-22 blockade or fasting impair intestinal homeostasis and restrict SFB expansion in mice.79 Work from Japan additionally revealed carbohydrates and insoluble fiber reduce IL-22 production and STAT3 phosphorylation in ILC3s, diminishing epithelial renewal and down-regulating the B3gnt7 gene.80 Western high-fat diets likewise impact SFB colonization. Such diets induce SFB loss in ILC3s-deficient, but not sufficient, mice, highlighting ILC3s as integral to high-fat diet mediated SFB depletion81 (Figure 4).
 |
Figure 4 SFB in ILC3s function or homeostasis. SFB colonise epithelial cells and initiate an IL-22-mediated Stat3 signalling cascade leading to local SAA1/2 expression and can also maintain the epithelial cell barrier function in the ileum by promoting the phosphorylation of STAT3 (pSTAT3) in the intestinal epithelium, inducing the production of REG3γ and contributing to IL-22 production by ILC3s. At the same time, ILC3s express high levels of Sca-1 on its surface in response to SFB colonisation, high levels of Sca-1 in response to SFB colonisation. Diet is also closely related to SFB function, as carbohydrates or insoluble cellulose fibre can affect IL-22 production by ILC3s and STAT3 phosphorylation in the ileum. |
Salmonella
Salmonella typhimurium is a parthenogenetic intracellular pathogen that invades and replicates within host cells, causing gastrointestinal and systemic disease.82,83 Salmonella establishes mucosal niches by inducing antimicrobial proteins that inhibit commensals while promoting its survival.84 During later infection stages, Salmonella targets ILC3s, activating IL-22 production through a TLR5-Myd88-IL-23 dependent mechanism enhancing APC-derived flagellin.85 Conversely, Salmonella directly induces caspase-1 mediated ILC3s cell death independently of flagellin.86 Casp1 ablation confers ILC3s survival and IL-22 elevation, exacerbating typhoid infection.86
ILC3s also shape the intestinal response to Salmonella via effects on epithelial fucosylation.87 While luminal colonization proves independent of fucosylation, epithelial fucose restricts bacterial tissue invasion, and its depletion elicits more severe infections.79
Akkermansia Muciniphila
Probiotics interact with intestinal immune cells and commensals to modulate immunity through health-promoting and immunomodulatory properties.88 Akkermansia muciniphila (A. muciniphila), a strictly anaerobic, mucin-degrading bacterium comprising 1–4% of the human gut microbiota, safeguards intestinal homeostasis via mucolytic actions.89–91 A. muciniphila promotes SCFAs (eg propionate, acetate) production and upregulates antimicrobial peptide genes (eg Reg3a, Reg3b, Reg3g), elevating paneth cell numbers yet exacerbating depletion following Salmonella typhimurium infection.92 A. muciniphila abundance modulates ILC3s-T cell balances, inhibits proinflammatory colonic epithelial signaling, and mitigates dysbiosis-driven inflammation93 (Figure 5).
 |
Figure 5 Probiotic Akkermansia muciniphila maintains intestinal barrier stability. Akkermansia muciniphila promotes the production of SCFA propionate and acetate, and upregulates gene expression of antimicrobial peptides (eg Reg3a, Reg3b and Reg3g). Co-colonisation with another bacterium, Salmonella typhimurium, exacerbated the reduction in cuprocytes, but co-colonisation with P. distasonis led to an increase in IL-17-ILC3s, reinforcing the interactions between gut microbes and their contribution to the protective immune response. |
The beneficial bacteria are Parabacteroides distasonis, Barnesiella intestinihominis in addition to Akkermansia muciniphila.94 A. muciniphila has been shown to enhance the number of ILC3s in the colon, a function that Amplification of this bacterium can be achieved through co-colonisation with Parabacteroides distasonis.89 Administration of A. muciniphila, either alone or in combination with P. distasonis, has been demonstrated to increase IL-17-ILC3s, thereby enhancing gut microbial interactions.93
These three types of gut microorganisms are all closely associated with human health. If their numbers and functions are not normalised, they will have a deleterious impact on human health. These gut flora are closely related to ILC3s, which can exert a direct or indirect influence on the equilibrium of the intestinal tract and the body’s overall health.
ILC3s in Autoimmune Disease
The function of ILC3s and their associated cytokines in maintaining intestinal immune homeostasis, as well as the impact of the intestinal microenvironment on ILC3s activity, have been previously described in detail. These processes may play a role in the pathogenesis of autoimmune diseases. Consequently, we have selected some representative autoimmune disorders for discussion in the following paragraphs.
IBD
Crohn’s disease (CD) and ulcerative colitis (UC) are classified as inflammatory bowel disease (IBD), a recurrent, non-specific lesion defined by chronic inflammation of the gastrointestinal system.95 A recent study employing single-cell sequencing to ascertain the transformation of immune cells in peripheral blood and colon tissue in patients with ulcerative colitis (UC) during active and remitting periods revealed a notable decline in the proportion of ILC3s.13 A reduction in the number of ILC3s, which produce IL-22, has been observed in the guts of patients with Crohn’s disease.96 Further analysis of animal studies revealed that ILC3s-deficient mice exhibited severe intestinal mucosal damage and developed colitis.97
A recent study has revealed that mice with colitis exhibited a notable elevation in aldosterone levels within the colon. Aldosterone plays a role in accelerating DSS-induced colitis, with ILC3s identified as the key effector cell. The activation of ILC3s by endogenous aldosterone resulted in the protection of mice from DSS-induced colitis. The halocorticoid receptor in ILC3s may play a pivotal role in responding to changes in aldosterone levels within the intestinal microenvironment, and in regulating IL17A stimulation of ILC3s.98
There is a correlation between IBD-related immunoinflammatory responses and imbalances in the gut microbiota.The imbalance in the effects of the gut microbiome on the intestinal mucosa and tissues is caused by an increase in the number of harmful bacteria (such as E. coli) and a decrease in the number of protective bacteria (such as bifidobacteria and Lactobacillus genera).99,100 Lactobacillus may promote intestinal mucous homeostasis by producing L-Orn (L-Ornithine). L-Orn stimulates Trp metabolism to produce AhR ligands in intestinal epithelial cells, thereby inducing the accumulation of ILC3s in intestinal tissues.3,101 A review of the literature revealed that the number of Lactobacillus with weaker attachments to epithelial cells colonizing the gut in individuals with IBD is less than that observed in healthy individuals.102 A reduction in the relative abundance of Lactobacillus, an increased severity of DSS-induced colitis, and an impairment of ILC3s development were observed.
Rheumatoid Arthritis
In the early stages of RA, the lymph node (LN) microenvironment is impaired, and activation and autoantibody production precede the onset of joint inflammation. It appears that ILCs in the LN microenvironment exhibit a shift from a more “homeostatic” to a more “inflammatory/activating” profile, characterised by an increased frequency of subpopulations of potentially pro-inflammatory cytokine-producing ILCs (ILC1s and ILC3s) in RA patients. It has been demonstrated that ILC1s and ILC3s are capable of inducing inflammation in RA pathogenesis.103,104 A clinical study revealed that ILC3s levels were elevated in RA patients and the frequency of ILC3s exhibited a correlation with the progression of RA. Additionally, the study demonstrated that ILC3s were positively correlated with Th1 and Th17 cells in peripheral blood in RA.14 A further study conducted in a Japanese RA population revealed a positive correlation between the incidence of ILC3s in synovial fluid and both tender joint counts and swollen joint counts.105 Furthermore, ILC3s were observed to be elevated in the synovium of CIA model mice in murine experiments. It is hypothesized that ILC3s may be involved in the pathogenesis of RA through the production of IL-17 and IL-22.105 ILC3s are a primary source of IL-22, IL-17 and TNFα.106 IL-22 plays a pivotal role in the inflammatory and proliferative cascade of various autoimmune diseases, including RA.107 IL-22 is also expressed as a pro-inflammatory cytokine in fibroblast-like synoviocytes (FLS) and macrophages, and promotes an inflammatory response on the FLS in patients with RA through the expression of IL-22 receptor 1. Other cytokines, such as TNFαand IL-17, also play an important role in synovitis in patients with RA.108 Although several clinical studies have indicated a potential association between ILC3s and RA exacerbations, the specific role of ILC3s in RA pathogenesis remains unclear.
A notable decline in the Firmicutes population was observed in RA patients, with a concomitant reduction in the production of butyrate-producing bacteria (BPB). Butyrate is a pivotal SCFA that exhibits a greater range of physiological functions than other SCFAs, such as acetate and propionate. The study observed an imbalance between butyrate producers and consumers in the gut microbiome of RA patients, which was found to be associated with levels of anti-citrullinated protein antibodies (ACPA) and joint deformities.109,110 The above demonstrates that SCFA is closely associated with ILC3s, which is also closely linked to RA. Consequently, we postulate that the intestinal flora in RA disease may have a certain relationship with ILC3s, which could contribute to the development of the disease.
Systemic Lupus Erythematosus
SLE is a heterogeneous autoimmune disease that predominantly affects women of childbearing age and can lead to damage in multiple organs.111,112 The immunopathogenesis of SLE involves various factors such as genetics, environment, hormones, epigenetics, and immune modulation. Autoreactive T cells play a pivotal role in the pathogenesis of SLE. Additionally, Th17 is implicated in the pathogenesis of SLE.113 ILC3s have been identified as the innate counterpart of Th17 cells.114 Clinical studies demonstrated alterations in the circulation of ILC in SLE patients with an increase in circulating ILC1s and ILC3s and a decrease in circulating ILC2s, indicating an imbalance in ILC homeostasis.15 Research by JIANG et al in 2020 found that SLE patients had significantly higher level of circulating ILC1s and ILC3s, with a significantly higher ILC1s/ILC3s ratio. In SLE patients with arthritis, there was a higher frequency of ILC3s and a lower ILC1s/ILC3s ratio, correlating with the presence of arthritis.115 Moreover, the absolute number of ILC3s was elevated in active SLE patients compared to those with inactive disease, and ILC3s levels correlated positively with disease activity and serum anti-dsDNA levels.115
Lupus nephritis (LN) is a serious complication of SLE that affects around 50% of SLE patients and significantly increases morbidity and mortality rates.116 Studies have shown that in LN patients, blood ILC2s levels are decreased, while ILC3s levels are significantly increased. Interestingly, the percentage of circulating ILC3s has been positively correlated with serum anti-double-stranded DNA (anti-dsDNA) antibodies and anti-nuclear antibodies (ANA). Research from mouse experiments has indicated that ILC3s from the intestinal can migrate to the kidneys. Additionally, it has been suggested that the CXCR6/CXCL16 pathway may play a crucial role in regulating the translocation of ILC3s from the intestine to the kidneys.117 These findings provide valuable insights into the potential involvement of ILC3s in the pathogenesis of SLE and LN. Further investigation into the mechanisms underlying the migration and function of ILC3s in LN may offer new therapeutic targets for the management of this severe complication of SLE.
Ankylosing Spondylitis
Ankylosing Spondylitis (AS) is a chronic inflammatory disease of unknown aetiology. In contrast to other systemic autoimmune diseases, the innate immune system plays a dominant role in AS. Aberrant activity of innate and innate-like immune cells, including ILC3s, has been observed in AS patients.118 It has been demonstrated that patients with AS exhibit a notable increase in IL-17-producing cells in the peripheral blood, inflamed bone marrow (BM), and synovial fluid. These cells also secrete high levels of IL-22 and α4β7 integrins, which are primarily responsible for directing immune cells to intestinal sites. In AS patients, The antireceptor of α4β7, MAdCAM1, is significantly upregulated in intestinal high endothelial venules (HEV) and BM in patients with AS, indicating its involvement in the chemotaxis of ILC3s to sites of active inflammation in AS.119–121 The correlation between ILC3s and AS disease activity scores suggests that ILC3s play a crucial role in AS pathogenesis and disease progression. The gut is regarded as a potential site of ILC3 differentiation, amplification and initiation in AS.122 Dysbiosis of the intestinal flora observed in AS patients may lead to the activation and proliferation of ILC3s, which could subsequently lead to joint inflammation and involvement.123
Other Autoimmune Diseases
A similar correlation has been observed in other immune diseases. To illustrate this point, consider the case of psoriasis, ILC3 has been identified in psoriatic skin samples from both humans and mice.124,125 Moreover, a decline in ILC3 numbers has been observed in response to therapeutic compounds, indicating their potential role in the pathogenesis of the condition. It has been demonstrated that tissue-resident ILCs, including ILC2, respond to IL-23 by activating a convergent, pathologically induced ILC3-like programme in Psoriasis. This programme is characterised by the production of cytokines such as IL-13 and IL-22, among other molecules.126
In patients diagnosed with type 1 diabetes (T1D), a reduced frequency of ILC3 was identified in the duodenum.127 In the pancreas and mesenteric lymph nodes of diabetic NOD mice, low IL-22-producing ILC3 was identified, alongside notably elevated levels of IFN-γ and TNF, but not IL-22. Furthermore, the transition from The transition from pre-diabetic to diabetic in NOD mice was associated with an impaired function of ILC3, which could lead to a reduction in the amount of Treg and suggest a protective effect of IL-22-producing and IL-22-producing ILC3 on T1D.128,129
Furthermore, in psoriatic arthritis, Soare et al observed that patients with psoriatic arthritis(PsA) exhibited elevated levels of circulating ILC3s and ILC2s, and that the ILC2/ILC3 ratio correlated with disease activity.87 Furthermore, Leijten et al demonstrated that ILC3s were amplified in the synovial fluid of patients with PsA, but not in peripheral blood.130 In ANCA-associated vasculitis (AAV), there is a reduction in the number of ILC2s and ILC3s, while the number of ILC1s is increased, in comparison to healthy controls or patients with remission-phase AAV. It is not yet possible to draw definitive conclusions regarding the role of ILCs in the pathogenesis of AAV.131
Discussion
Recent research underscores the pivotal role of ILC3s in modulating innate immune responses. These cells are ubiquitously distributed across the body, with a constant presence at mucosal barrier locales such as the lungs, liver, intestines, spleen, skin, and secondary lymphoid tissues.132 Circulating ILCs progenitors, originating from the fetal liver, migrate to the bone marrow (BM) and persist through to adulthood.133 Primarily, ILC3s are drawn to intestinal tissues, moving from the bone marrow to mesenteric lymph nodes in a CCR7-dependent manner, with commensal bacteria potentially directing intestinal ILC3s towards the lungs.133 The lung-specific migration of ILC3s involves CCR4 and CXCR5 signaling pathways, where CCR4 acts as a homing receptor and CXCR5 signaling is essential for lung colonization and subsequent IL-17 production, orchestrating neutrophilic responses.134,135 Additionally, ILC3s can be summoned to inflamed skin via a CCR6-dependent route, influencing the equilibrium of commensalism by modulating sebocyte functionality through direct interactions with CD4 T cells.136 Within the skin, ILC3s secrete IL-22 to bolster antimicrobial responses by keratinocytes, whereas IL-17 prompts keratinocytes to release IL-33, intensifying inflammation and fostering type 2 immune reactions.137
In the blood a considerable number of ILCs progenitors exist, yet mature ILC1s and ILC3s are notably absent.133 Recent discoveries have unveiled an unrecognized ILCs subset within the liver, exhibiting an ILC3s-like phenotype with the capability to produce the ILC2s-specific cytokine IL-13.138 Notably, variations in cytokines, such as IL-13, IL-17 and IL-22 across different tissues (intestines, lungs, skin), present substantial challenges in the consistency and correlation of research findings across diverse populations, necessitating further investigative efforts.
In vivo, ILC3s are integral to a myriad of protective homeostatic functions. However, imbalances in ILC3s responses are linked with a range of chronic inflammatory ailments and are progressively acknowledged for their role in the emergence and evolution of autoimmune diseases, including SLE,15,115 AS,123 psoriasis,126 type 1 diabetes,127 Rheumatoid Arthritis.107,108 The precise aetiology and pathogenesis of these autoimmune diseases in relation to ILC3s remain unclear. Despite the existence of clinical evidence suggesting a correlation between ILC3s and the aforementioned autoimmune diseases, there is a distinct lack of scientific literature that clarifies the specific interactions between ILC3s and them. Furthermore, a plethora of autoimmune diseases, including vitiligo, pernicious anaemia, have not yet been clinically linked to ILC3s, thus representing a relatively novel perspective that merits rigorous study and exploration.
The microenvironment exerts a substantial influence on immune cell plasticity, with cytokines and transcription factors playing a pivotal role in the differentiation and maintenance of ILCs. These factors also influence the distribution of ILCs subpopulations, which may consequently alter the trajectory of the disease. Current data can suggest that ILCs can be utilised as biomarkers to assess disease severity or treatment response. Recent technological advances have enhanced our capacity to monitor and characterise ILC3s dynamics in the gut microenvironment, thereby facilitating the development of personalised therapeutic strategies. Single-cell RNA sequencing (scRNA-seq) and mass spectrometry cell counting (CyTOF) now enable high-resolution analyses of ILC3s heterogeneity, unveiling subtype-specific functional states in autoimmune environments.13,139,140 The following experiment will utilise spatial transcriptomics and multiphoton imaging to track ILC3s interactions with gut microbiota and epithelial cells in real time in a living organism.141,142 These innovations have the potential to assist in the stratification of patients according to ILC3s characteristics, thus facilitating the development of targeted interventions such as cytokine modulation (eg anti-IL-23 therapy) or microbiota-directed therapies. However, the standardisation of these technologies in clinical applications remains challenging, particularly with regard to cost-effectiveness and longitudinal monitoring. The development of organoid-ILC3s co-culture models in conjunction with AI-driven biomarker discovery may serve to bridge this translational gap in the future.
Data Sharing Statement
No data was used for the research described in the article.
Funding
This research was supported by National Natural Science Foundation of China (No. 82274382, 82405212), Natural Science Foundation of Zhejiang Province (LY24H270004), Postdoctoral Fellowship Program of CPSF(GZC20232374).
Disclosure
The authors declare no conflict of interest.
References
1. Gelmez MY, Cinar S, Cetin EA. et al. Inflammatory status might direct ILC and NK cells to IL-17 expressing ILC3 and NK subsets in Behcet’s disease. Immunol Lett. 2021;235:1–8. doi:10.1016/j.imlet.2021.04.008
2. Zeng B, Shi S, Ashworth G, et al. ILC3 function as a double-edged sword in inflammatory bowel diseases. Cell Death Dis. 2019;10(4):315. doi:10.1038/s41419-019-1540-2
3. Zhong C, Zheng M, Cui K, et al. Differential expression of the transcription factor gata3 specifies lineage and functions of innate lymphoid cells. Immunity. 2020;52(1):83–95.e84. doi:10.1016/j.immuni.2019.12.001
4. Siggs OM, Li X, Xia Y, et al. ZBTB1 is a determinant of lymphoid development. J Exp Med. 2012;209(1):19–27. doi:10.1084/jem.20112084
5. Masoud GN, Li W. HIF-1α pathway: role, regulation and intervention for cancer therapy. Acta Pharm Sin B. 2015;5(5):378–389. doi:10.1016/j.apsb.2015.05.007
6. Spits H, Di Santo JP. The expanding family of innate lymphoid cells: regulators and effectors of immunity and tissue remodeling. Nat Immunol. 2011;12(1):21–27. doi:10.1038/ni.1962
7. Spits H, Artis D, Colonna M, et al. Innate lymphoid cells--a proposal for uniform nomenclature. Nat Rev Immunol. 2013;13(2):145–149. doi:10.1038/nri3365
8. Qi H, Li Y, Yun H, et al. Lactobacillus maintains healthy gut mucosa by producing L-Ornithine. Commun Biol. 2019;2(1):171. doi:10.1038/s42003-019-0424-4
9. Sah P, Zenewicz LA. The polyamine putrescine is a positive regulator of group 3 innate lymphocyte activation. Immunohorizons. 2023;7(1):41–48. doi:10.4049/immunohorizons.2200097
10. Jarade A, Garcia Z, Marie S, et al. Inflammation triggers ILC3 patrolling of the intestinal barrier. Nat Immunol. 2022;23(9):1317–1323. doi:10.1038/s41590-022-01284-1
11. Guo X, Liang Y, Zhang Y, et al. Innate lymphoid cells control early colonization resistance against intestinal pathogens through ID2-dependent regulation of the microbiota. Immunity. 2015;42(4):731–743. doi:10.1016/j.immuni.2015.03.012
12. Serafini N, Jarade A, Surace L, et al. Trained ILC3 responses promote intestinal defense. Science. 2022;375(6583):859–863. doi:10.1126/science.aaz8777
13. Mitsialis V, Wall S, Liu P, et al. Single-cell analyses of colon and blood reveal distinct immune cell signatures of ulcerative colitis and Crohn’s disease. Gastroenterology. 2020;159(2):591–608.e510. doi:10.1053/j.gastro.2020.04.074
14. Liu X, Wu L, Chen S, et al. Group 3 innate lymphoid cells promotes Th17 cells differentiation in rheumatoid arthritis. Clin Exp Rheumatol. 2024. doi:10.55563/clinexprheumatol/ktbiqc
15. Hou M, Liu S. Innate lymphoid cells are increased in systemic lupus erythematosus. Clin Exp Rheumatol. 2019;37(4):676–679.
16. Sasaki T, Moro K, Kubota T, et al. Innate lymphoid cells in the induction of obesity. Cell Rep. 2019;28(1):202–217.e207. doi:10.1016/j.celrep.2019.06.016
17. Vivier E, Artis D, Colonna M, et al. Innate lymphoid cells: 10 years on. Cell. 2018;174(5):1054–1066. doi:10.1016/j.cell.2018.07.017
18. Lv X, Zhu S, Wu J, et al. Reciprocal costimulatory molecules control the activation of mucosal type 3 innate lymphoid cells during engagement with B cells. Cell mol Immunol. 2023;20(7):808–819. doi:10.1038/s41423-023-01041-w
19. Steffen J, Ehrentraut S, Bank U, et al. Type 1 innate lymphoid cells regulate the onset of Toxoplasma gondii-induced neuroinflammation. Cell Rep. 2022;38(13):110564. doi:10.1016/j.celrep.2022.110564
20. Klose CSN, Flach M, Möhle L, et al. Differentiation of type 1 ILCs from a common progenitor to all helper-like innate lymphoid cell lineages. Cell. 2014;157(2):340–356. doi:10.1016/j.cell.2014.03.030
21. Daussy C, Faure F, Mayol K, et al. T-bet and Eomes instruct the development of two distinct natural killer cell lineages in the liver and in the bone marrow. J Exp Med. 2014;211(3):563–577. doi:10.1084/jem.20131560
22. Kawano R, Okamura T, Hashimoto Y, et al. Erythritol ameliorates small intestinal inflammation induced by high-fat diets and improves glucose tolerance. Int J mol Sci. 2021;22(11):5558. doi:10.3390/ijms22115558
23. Zhong C, Zheng M, Zhu J. Lymphoid tissue inducer-A divergent member of the ILC family. Cytokine Growth Factor Rev. 2018;42:5–12. doi:10.1016/j.cytogfr.2018.02.004
24. Bando JK, Liang HE, Locksley RM. Identification and distribution of developing innate lymphoid cells in the fetal mouse intestine. Nat Immunol. 2015;16(2):153–160. doi:10.1038/ni.3057
25. Rethacker L, Boy M, Bisio V, et al. Innate lymphoid cells: NK and cytotoxic ILC3 subsets infiltrate metastatic breast cancer lymph nodes. Oncoimmunology. 2022;11(1):2057396. doi:10.1080/2162402x.2022.2057396
26. Melo-Gonzalez F, Hepworth MR. Functional and phenotypic heterogeneity of group 3 innate lymphoid cells. Immunology. 2017;150(3):265–275. doi:10.1111/imm.12697
27. Hoorweg K, Peters CP, Cornelissen F, et al. Functional differences between human NKp44(-) and NKp44(+) RORC(+) innate lymphoid cells. Front Immunol. 2012;3:72. doi:10.3389/fimmu.2012.00072
28. Klose CS, Kiss EA, Schwierzeck V, et al. A T-bet gradient controls the fate and function of CCR6-RORγt+ innate lymphoid cells. Nature. 2013;494(7436):261–265. doi:10.1038/nature11813
29. Zheng M, Mao K, Fang D, et al. B cell residency but not T cell-independent IgA switching in the gut requires innate lymphoid cells. Proc Natl Acad Sci U S A. 2021;118(27). doi:10.1073/pnas.2106754118
30. Eberl G, Marmon S, Sunshine MJ, et al. An essential function for the nuclear receptor RORgamma(t) in the generation of fetal lymphoid tissue inducer cells. Nat Immunol. 2004;5(1):64–73. doi:10.1038/ni1022
31. Lu Y, Zhang X, Bouladoux N, et al. Zbtb1 controls NKp46(+) ROR-gamma-T(+) innate lymphoid cell (ILC3) development. Oncotarget. 2017;8(34):55877–55888. doi:10.18632/oncotarget.19645
32. Xu W, Cherrier DE, Chea S, et al. An Id2(RFP)-reporter mouse redefines innate lymphoid cell precursor potentials. Immunity. 2019;50(4):1054–1068.e1053. doi:10.1016/j.immuni.2019.02.022
33. Serafini N, Klein wolterink RG, Satoh-Takayama N, et al. Gata3 drives development of RORγt+ group 3 innate lymphoid cells. J Exp Med. 2014;211(2):199–208. doi:10.1084/jem.20131038
34. Zhong C, Cui K, Wilhelm C, et al. Group 3 innate lymphoid cells continuously require the transcription factor GATA-3 after commitment. Nat Immunol. 2016;17(2):169–178. doi:10.1038/ni.3318
35. Yagi R, Zhong C, Northrup DL, et al. The transcription factor GATA3 is critical for the development of all IL-7Rα-expressing innate lymphoid cells. Immunity. 2014;40(3):378–388. doi:10.1016/j.immuni.2014.01.012
36. Lo BC, Gold MJ, Hughes MR, et al. The orphan nuclear receptor ROR alpha and group 3 innate lymphoid cells drive fibrosis in a mouse model of Crohn’s disease. Sci Immunol. 2016;1(3). doi:10.1126/sciimmunol.aaf8864
37. Seillet C, Rankin LC, Groom JR, et al. Nfil3 is required for the development of all innate lymphoid cell subsets. J Exp Med. 2014;211(9):1733–1740. doi:10.1084/jem.20140145
38. Geiger TL, Abt MC, Gasteiger G, et al. Nfil3 is crucial for development of innate lymphoid cells and host protection against intestinal pathogens. J Exp Med. 2014;211(9):1723–1731. doi:10.1084/jem.20140212
39. Liu Q, Kim MH, Friesen L, et al. BATF regulates innate lymphoid cell hematopoiesis and homeostasis. Sci Immunol. 2020;5(54). doi:10.1126/sciimmunol.aaz8154
40. Kim H, Dejsuphong D, Adelmant G, et al. Transcriptional repressor ZBTB1 promotes chromatin remodeling and translesion DNA synthesis. Mol Cell. 2014;54(1):107–118. doi:10.1016/j.molcel.2014.02.017
41. Abidi A, Laurent T, Bériou G, et al. Characterization of Rat ILCs Reveals ILC2 as the dominant intestinal subset. Front Immunol. 2020;11:255. doi:10.3389/fimmu.2020.00255
42. Fachi JL, Pral LP, Dos Santos JAC, et al. Hypoxia enhances ILC3 responses through HIF-1α-dependent mechanism. Mucosal Immunol. 2021;14(4):828–841. doi:10.1038/s41385-020-00371-6
43. Fachi JL, Pral LP, Assis HC, et al. Hyperbaric oxygen augments susceptibility to C. difficile infection by impairing gut microbiota ability to stimulate the HIF-1α-IL-22 axis in ILC3. Gut Microbes. 2024;16(1):2297872. doi:10.1080/19490976.2023.2297872
44. Krzywinska E, Sobecki M, Nagarajan S, et al. The transcription factor HIF-1α mediates plasticity of NKp46+ innate lymphoid cells in the gut. J Exp Med. 2022;219(2). doi:10.1084/jem.20210909
45. Ciofani M, Madar A, Galan C, et al. A validated regulatory network for Th17 cell specification. Cell. 2012;151(2):289–303. doi:10.1016/j.cell.2012.09.016
46. Gury-BenAri M, Thaiss CA, Serafini N, et al. The spectrum and regulatory landscape of intestinal innate lymphoid cells are shaped by the microbiome. Cell. 2016;166(5):1231–1246.e1213. doi:10.1016/j.cell.2016.07.043
47. Parker ME, Barrera A, Wheaton JD, et al. c-Maf regulates the plasticity of group 3 innate lymphoid cells by restraining the type 1 program. J Exp Med. 2020;217(1). doi:10.1084/jem.20191030
48. Pokrovskii M, Hall JA, Ochayon DE, et al. Characterization of transcriptional regulatory networks that promote and restrict identities and functions of intestinal innate lymphoid cells. Immunity. 2019;51(1):185–197.e186. doi:10.1016/j.immuni.2019.06.001
49. Tizian C, Lahmann A, Hölsken O, et al. c-Maf restrains T-bet-driven programming of CCR6-negative group 3 innate lymphoid cells. Elife. 2020;9. doi:10.7554/eLife.52549
50. Sonnenberg GF, Monticelli LA, Alenghat T, et al. Innate lymphoid cells promote anatomical containment of lymphoid-resident commensal bacteria. Science. 2012;336(6086):1321–1325. doi:10.1126/science.1222551
51. Sonnenberg GF, Artis D. Innate lymphoid cells in the initiation, regulation and resolution of inflammation. Nat Med. 2015;21(7):698–708. doi:10.1038/nm.3892
52. Tan JK, McKenzie C, Mariño E, et al. Metabolite-sensing G protein-coupled receptors-facilitators of diet-related immune regulation. Annu Rev Immunol. 2017;35(1):371–402. doi:10.1146/annurev-immunol-051116-052235
53. Zhou J, Yue J, Yao Y, et al. Dihydromyricetin protects intestinal barrier integrity by promoting IL-22 expression in ILC3s through the AMPK/SIRT3/STAT3 signaling pathway. Nutrients. 2023;15. doi:10.3390/nu15020355
54. Deng F, Hu JJ, Lin ZB, et al. Gut microbe-derived milnacipran enhances tolerance to gut ischemia/reperfusion injury. Cell Rep Med. 2023;4(3):100979. doi:10.1016/j.xcrm.2023.100979
55. Sondermann NC, Faßbender S, Hartung F, et al. Functions of the aryl hydrocarbon receptor (AHR) beyond the canonical AHR/ARNT signaling pathway. Biochem Pharmacol. 2023;208:115371. doi:10.1016/j.bcp.2022.115371
56. Xia Y, Liu C, Li R, et al. Lactobacillus-Derived Indole-3-Lactic Acid ameliorates Colitis in Cesarean-Born Offspring via Activation of Aryl Hydrocarbon Receptor. iScience 26.2023.108279 doi:10.1016/j.isci.2023.108279
57. Shi Z, Ohno H, Satoh-Takayama N. Dietary derived micronutrients modulate immune responses through innate lymphoid cells. Front Immunol. 2021;12:670632. doi:10.3389/fimmu.2021.670632
58. Xia P, Liu J, Wang S, et al. WASH maintains NKp46(+) ILC3 cells by promoting AHR expression. Nat Commun. 2017;8(1):15685. doi:10.1038/ncomms15685
59. Tan C, Hong G, Wang Z, et al. Promoting effect of l-fucose on the regeneration of intestinal stem cells through AHR/IL-22 pathway of intestinal lamina propria monocytes. Nutrients. 2022;14(22):4789. doi:10.3390/nu14224789
60. Chun E, Lavoie S, Fonseca-Pereira D, et al. Metabolite-sensing receptor Ffar2 regulates colonic group 3 innate lymphoid cells and gut immunity. Immunity. 2019;51(5):871–884.e876. doi:10.1016/j.immuni.2019.09.014
61. Sepahi A, Liu Q, Friesen L, et al. Dietary fiber metabolites regulate innate lymphoid cell responses. Mucosal Immunol. 2021;14(2):317–330. doi:10.1038/s41385-020-0312-8
62. Alisjahbana A, Willinger T. Metabolite sensing by colonic ILC3s: how far is too Ffar2 go? Immunity. 2019;51(5):786–788. doi:10.1016/j.immuni.2019.10.005
63. Willinger T. Metabolic control of innate lymphoid cell migration. Front Immunol. 2019;10:2010. doi:10.3389/fimmu.2019.02010
64. Kim MH, Taparowsky EJ, Kim CH. Retinoic acid differentially regulates the migration of innate lymphoid cell subsets to the gut. Immunity. 2015;43(1):107–119. doi:10.1016/j.immuni.2015.06.009
65. Ruiter B, Patil SU, Shreffler WG. Vitamins A and D have antagonistic effects on expression of effector cytokines and gut-homing integrin in human innate lymphoid cells. Clin Exp Allergy. 2015;45(7):1214–1225. doi:10.1111/cea.12568
66. Mielke LA, Jones SA, Raverdeau M, et al. Retinoic acid expression associates with enhanced IL-22 production by γδ T cells and innate lymphoid cells and attenuation of intestinal inflammation. J Exp Med. 2013;210(6):1117–1124. doi:10.1084/jem.20121588
67. Fachi JL, Sécca C, Rodrigues PB, et al. Acetate coordinates neutrophil and ILC3 responses against C. difficile through FFAR2. J Exp Med. 2020;217(3). doi:10.1084/jem.20190489
68. Kim SH, Cho BH, Kiyono H, et al. Microbiota-derived butyrate suppresses group 3 innate lymphoid cells in terminal ileal Peyer’s patches. Sci Rep. 2017;7(1):3980. doi:10.1038/s41598-017-02729-6
69. Drommi F, Calabrò A, Vento G, et al. Crosstalk between ILC3s and microbiota: implications for colon cancer development and treatment with immune check point inhibitors. Cancers (Basel). 2023;15(11):2893. doi:10.3390/cancers15112893
70. Goverse G, Labao-Almeida C, Ferreira M, et al. Vitamin A controls the presence of RORγ+ innate lymphoid cells and lymphoid tissue in the small intestine. J Immunol. 2016;196(12):5148–5155. doi:10.4049/jimmunol.1501106
71. Noh S, Park CO, Bae JM, et al. Lower vitamin D status is closely correlated with eczema of the head and neck. J Allergy Clin Immunol. 2014;133(6):1767–1770.e1766. doi:10.1016/j.jaci.2014.02.038
72. He L, Zhou M, Li YC. Vitamin D/Vitamin D receptor signaling is required for normal development and function of group 3 innate lymphoid cells in the gut. iScience. 2019;17:119–131. doi:10.1016/j.isci.2019.06.026
73. Lin YD, Arora J, Diehl K, et al. Vitamin D is required for ILC3 Derived IL-22 and protection from citrobacter rodentium infection. Front Immunol. 2019;10:1. doi:10.3389/fimmu.2019.00001
74. Klaasen HL, Koopman JP, Van den Brink ME, et al. Intestinal, segmented, filamentous bacteria in a wide range of vertebrate species. Lab Anim. 1993;27(2):141–150. doi:10.1258/002367793780810441
75. Eckhardt ER, Witta J, Zhong J, et al. Intestinal epithelial serum amyloid A modulates bacterial growth in vitro and pro-inflammatory responses in mouse experimental colitis. BMC Gastroenterol. 2010;10(1):133. doi:10.1186/1471-230x-10-133
76. Rutz S, Wang X, Ouyang W. The IL-20 subfamily of cytokines--from host defence to tissue homeostasis. Nat Rev Immunol. 2014;14(12):783–795. doi:10.1038/nri3766
77. Sano T, Huang W, Hall JA, et al. An IL-23R/IL-22 circuit regulates epithelial serum amyloid A to promote local effector Th17 responses. Cell. 2016;164(1–2):324. doi:10.1016/j.cell.2015.12.047
78. Brooks JF 2nd, Behrendt CL, Ruhn KA, et al. The microbiota coordinates diurnal rhythms in innate immunity with the circadian clock. Cell. 2021;184(16):4154–4167.e4112. doi:10.1016/j.cell.2021.07.001
79. Goto Y, Obata T, Kunisawa J, et al. Innate lymphoid cells regulate intestinal epithelial cell glycosylation. Science. 2014;345(6202):1254009. doi:10.1126/science.1254009
80. Pellizzon M. Choice of laboratory animal diet influences intestinal health. Lab anim NY. 2016;45(6):238–239. doi:10.1038/laban.1014
81. Kawano Y, Edwards M, Huang Y, et al. Microbiota imbalance induced by dietary sugar disrupts immune-mediated protection from metabolic syndrome. Cell. 2022;185(19):3501–3519.e3520. doi:10.1016/j.cell.2022.08.005
82. Chong A, Starr T, Finn CE, et al. A role for the salmonella type III secretion system 1 in bacterial adaptation to the cytosol of epithelial cells. Mol Microbiol. 2019;112(4):1270–1283. doi:10.1111/mmi.14361
83. Jennings E, Thurston TLM, Holden DW. Salmonella SPI-2 Type III secretion system effectors: molecular mechanisms and physiological consequences. Cell Host Microbe. 2017;22(2):217–231. doi:10.1016/j.chom.2017.07.009
84. Grizotte-Lake M, Zhong G, Duncan K, et al. Commensals suppress intestinal epithelial cell retinoic acid synthesis to regulate interleukin-22 activity and prevent microbial dysbiosis. Immunity. 2018;49(6):1103–1115.e1106. doi:10.1016/j.immuni.2018.11.018
85. Xiong L, Wang S, Dean JW, et al. Group 3 innate lymphoid cell pyroptosis represents a host defence mechanism against Salmonella infection. Nat Microbiol. 2022;7(7):1087–1099. doi:10.1038/s41564-022-01142-8
86. Joseph AM, Sonnenberg GF. ILC3 pyroptosis limits Salmonella infection. Nat Microbiol. 2022;7(7):933–934. doi:10.1038/s41564-022-01165-1
87. Soare A, Weber S, Maul L, et al. Cutting edge: homeostasis of innate lymphoid cells is imbalanced in psoriatic arthritis. J Immunol. 2018;200(4):1249–1254. doi:10.4049/jimmunol.1700596
88. Mazziotta C, Tognon M, Martini F, et al. Probiotics mechanism of action on immune cells and beneficial effects on human health. Cells. 2023;12(1):184. doi:10.3390/cells12010184
89. Paone P, Cani PD. Mucus barrier, mucins and gut microbiota: the expected slimy partners? Gut. 2020;69(12):2232–2243. doi:10.1136/gutjnl-2020-322260
90. Derrien M, Collado MC, Ben-Amor K, et al. The mucin degrader akkermansia muciniphila is an abundant resident of the human intestinal tract. Appl Environ Microbiol. 2008;74(5):1646–1648. doi:10.1128/aem.01226-07
91. Ganesh BP, Klopfleisch R, Loh G, et al. Commensal Akkermansia muciniphila exacerbates gut inflammation in Salmonella Typhimurium-infected gnotobiotic mice. PLoS One. 2013;8(9):e74963. doi:10.1371/journal.pone.0074963
92. Han Y, Ling Q, Wu L, et al. Akkermansia muciniphila inhibits nonalcoholic steatohepatitis by orchestrating TLR2-activated γδT17 cell and macrophage polarization. Gut Microbes. 2023;15(1):2221485. doi:10.1080/19490976.2023.2221485
93. Gaifem J, Mendes-Frias A, Wolter M, et al. Akkermansia muciniphila and Parabacteroides distasonis synergistically protect from colitis by promoting ILC3 in the gut. mBio. 2024;15(4):e0007824. doi:10.1128/mbio.00078-24
94. Bai X, Fu R, Liu Y, et al. Ginsenoside Rk3 modulates gut microbiota and regulates immune response of group 3 innate lymphoid cells to against colorectal tumorigenesis. J Pharm Anal. 2024;14(2):259–275. doi:10.1016/j.jpha.2023.09.010
95. Ye Q, Huang S, Wang Y, et al. Wogonin improves colitis by activating the AhR pathway to regulate the plasticity of ILC3/ILC1. Phytomedicine. 2024;128:155425. doi:10.1016/j.phymed.2024.155425
96. Li J, Doty AL, Tang Y, et al. Enrichment of IL-17A(+) IFN-γ(+) and IL-22(+) IFN-γ(+) T cell subsets is associated with reduction of NKp44(+) ILC3s in the terminal ileum of Crohn’s disease patients. Clin Exp Immunol. 2017;190(1):143–153. doi:10.1111/cei.12996
97. Aparicio-Domingo P, Romera-Hernandez M, Karrich JJ, et al. Type 3 innate lymphoid cells maintain intestinal epithelial stem cells after tissue damage. J Exp Med. 2015;212(11):1783–1791. doi:10.1084/jem.20150318
98. Zhao R, Hong L, Shi G, et al. Mineralocorticoid promotes intestinal inflammation through receptor dependent IL17 production in ILC3s. Int Immunopharmacol. 2024;130:111678. doi:10.1016/j.intimp.2024.111678
99. Ng SC, Hart AL, Kamm MA, et al. Mechanisms of action of probiotics: recent advances. Inflamm Bowel Dis. 2009;15(2):300–310. doi:10.1002/ibd.20602
100. Dominguez-Bello MG, De jesus-laboy KM, Shen N, et al. Partial restoration of the microbiota of cesarean-born infants via vaginal microbial transfer. Nat Med. 2016;22(3):250–253. doi:10.1038/nm.4039
101. Dominguez-Bello MG, Costello EK, Contreras M, et al. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc Natl Acad Sci U S A. 2010;107(26):11971–11975. doi:10.1073/pnas.1002601107
102. Najafi S, Sotoodehnejadnematalahi F, Amiri MM, et al. Decreased mucosal adhesion of Lactobacillus species in patients with inflammatory bowel disease. Caspian J Intern Med. 2022;13(4):713–720. doi:10.22088/cjim.13.4.71
103. Rodríguez-Carrio J, Hähnlein JS, Ramwadhdoebe TH, et al. Brief report: altered innate lymphoid cell subsets in human lymph node biopsy specimens obtained during the at-risk and earliest phases of rheumatoid arthritis. Arthritis Rheumatol. 2017;69(1):70–76. doi:10.1002/art.39811
104. Dalbeth N, Callan MF. A subset of natural killer cells is greatly expanded within inflamed joints. Arthritis Rheum. 2002;46(7):1763–1772. doi:10.1002/art.10410
105. Takaki-Kuwahara A, Arinobu Y, Miyawaki K, et al. CCR6+ group 3 innate lymphoid cells accumulate in inflamed joints in rheumatoid arthritis and produce Th17 cytokines. Arthritis Res Ther. 2019;21(1):198. doi:10.1186/s13075-019-1984-x
106. Ren J, Feng Z, Lv Z, et al. Natural killer-22 cells in the synovial fluid of patients with rheumatoid arthritis are an innate source of interleukin 22 and tumor necrosis factor-α. J Rheumatol. 2011;38(10):2112–2118. doi:10.3899/jrheum.101377
107. Ciccia F, Accardo-Palumbo A, Alessandro R, et al. Interleukin-22 and interleukin-22-producing NKp44+ natural killer cells in subclinical gut inflammation in ankylosing spondylitis. Arthritis Rheum. 2012;64(6):1869–1878. doi:10.1002/art.34355
108. van Hamburg JP, Corneth OB, Paulissen SM, et al. IL-17/Th17 mediated synovial inflammation is IL-22 independent. Ann Rheum Dis. 2013;72(10):1700–1707. doi:10.1136/annrheumdis-2012-202373
109. Kishikawa T, Maeda Y, Nii T, et al. Metagenome-wide association study of gut microbiome revealed novel aetiology of rheumatoid arthritis in the Japanese population. Ann Rheum Dis. 2020;79(1):103–111. doi:10.1136/annrheumdis-2019-215743
110. Wang H, Cai Y, Wu W, et al. Exploring the role of gut microbiome in autoimmune diseases: a comprehensive review. Autoimmun Rev. 2024;23(12):103654. doi:10.1016/j.autrev.2024.103654
111. Tsokos GC. Systemic lupus erythematosus. N Engl J Med. 2011;365(22):2110–2121. doi:10.1056/NEJMra1100359
112. Zhang Y, Du L, Wang C, et al. Neddylation is a novel therapeutic target for lupus by regulating double negative T cell homeostasis. Signal Transduct Target Ther. 2024;9(1):18. doi:10.1038/s41392-023-01709-9
113. Paroli M, Caccavale R, Fiorillo MT, et al. The double game played by Th17 cells in infection: host defense and immunopathology. Pathogens. 2022;11(12):1547. doi:10.3390/pathogens11121547
114. Peng V, Cao S, Trsan T, et al. Ornithine decarboxylase supports ILC3 responses in infectious and autoimmune colitis through positive regulation of IL-22 transcription. Proc Natl Acad Sci U S A. 2022;119(45):e2214900119. doi:10.1073/pnas.2214900119
115. Jiang Y, Zhao Y, Liu Y, et al. Imbalanced innate lymphoid cells are associated with disease activity and arthritis involvement in patients with systemic lupus erythematosus. Arch Rheumatol. 2020;35(4):521–532. doi:10.46497/ArchRheumatol.2020.7440
116. Kiriakidou M, Ching CL. Systemic lupus erythematosus. Ann Intern Med. 2020;172(11):Itc81–itc96. doi:10.7326/aitc202006020
117. Li F, Liang Z, Zhong H, et al. Group 3 innate lymphoid cells exacerbate lupus nephritis by promoting B cell activation in kidney ectopic lymphoid structures. Adv Sci (Weinh). 2023;10(35):e2302804. doi:10.1002/advs.202302804
118. Mauro D, Thomas R, Guggino G, et al. Ankylosing spondylitis: an autoimmune or autoinflammatory disease? Nat Rev Rheumatol. 2021;17(7):387–404. doi:10.1038/s41584-021-00625-y
119. Mora JR, Bono MR, Manjunath N, et al. Selective imprinting of gut-homing T cells by Peyer’s patch dendritic cells. Nature. 2003;424(6944):88–93. doi:10.1038/nature01726
120. Berlin C, Berg EL, Briskin MJ, et al. Alpha 4 beta 7 integrin mediates lymphocyte binding to the mucosal vascular addressin MAdCAM-1. Cell. 1993;74(1):185–195. doi:10.1016/0092-8674(93)90305-a
121. Ciccia F, Guggino G, Rizzo A, et al. Type 3 innate lymphoid cells producing IL-17 and IL-22 are expanded in the gut, in the peripheral blood, synovial fluid and bone marrow of patients with ankylosing spondylitis. Ann Rheum Dis. 2015;74(9):1739–1747. doi:10.1136/annrheumdis-2014-206323
122. Neerinckx B, Elewaut D, Lories RJ. Spreading spondyloarthritis: are ILCs cytokine shuttles from base camp gut? Ann Rheum Dis. 2015;74(9):1633–1635. doi:10.1136/annrheumdis-2015-207735
123. Ruutu M, Thomas G, Steck R, et al. β-glucan triggers spondylarthritis and Crohn’s disease-like ileitis in SKG mice. Arthritis Rheum. 2012;64(7):2211–2222. doi:10.1002/art.34423
124. Teunissen MBM, Munneke JM, Bernink JH, et al. Composition of innate lymphoid cell subsets in the human skin: enrichment of NCR(+) ILC3 in lesional skin and blood of psoriasis patients. J Invest Dermatol. 2014;134(9):2351–2360. doi:10.1038/jid.2014.146
125. Villanova F, Flutter B, Tosi I, et al. Characterization of innate lymphoid cells in human skin and blood demonstrates increase of NKp44+ ILC3 in psoriasis. J Invest Dermatol. 2014;134(4):984–991. doi:10.1038/jid.2013.477
126. Bielecki P, Riesenfeld SJ, Hütter JC, et al. Skin-resident innate lymphoid cells converge on a pathogenic effector state. Nature. 2021;592(7852):128–132. doi:10.1038/s41586-021-03188-w
127. Graves CL, Li J, LaPato M, et al. Intestinal epithelial cell regulation of adaptive immune dysfunction in human type 1 diabetes. Front Immunol. 2016;7:679. doi:10.3389/fimmu.2016.00679
128. Miani M, Le Naour J, Waeckel-Enée E, et al. Gut microbiota-stimulated innate lymphoid cells support β-defensin 14 expression in pancreatic endocrine cells, preventing autoimmune diabetes. Cell Metab. 2018;28(4):557–572.e556. doi:10.1016/j.cmet.2018.06.012
129. Saksida T, Paunović V, Koprivica I, et al. Development of type 1 diabetes in mice is associated with a decrease in IL-2-Producing ILC3 and FoxP3(+) treg in the small intestine. Molecules. 2023;28(8):3366. doi:10.3390/molecules28083366
130. Leijten EF, van Kempen TS, Boes M, et al. Brief report: enrichment of activated group 3 innate lymphoid cells in psoriatic arthritis synovial fluid. Arthritis Rheumatol. 2015;67(10):2673–2678. doi:10.1002/art.39261
131. Braudeau C, Amouriaux K, Néel A, et al. Persistent deficiency of circulating mucosal-associated invariant T (MAIT) cells in ANCA-associated vasculitis. J Autoimmun. 2016;70:73–79. doi:10.1016/j.jaut.2016.03.015
132. Hernández-Torres DC, Stehle C. Embryonic ILC-poiesis across tissues. Front Immunol. 2022;13:1040624. doi:10.3389/fimmu.2022.1040624
133. Zhu Q, Gao P, Tober J, et al. Developmental trajectory of prehematopoietic stem cell formation from endothelium. Blood. 2020;136(7):845–856. doi:10.1182/blood.2020004801
134. Das S, Chauhan KS, Ahmed M, et al. Lung type 3 innate lymphoid cells respond early following Mycobacterium tuberculosis infection. mBio. 2024;15(4):e0329923. doi:10.1128/mbio.03299-23
135. Zhang K, Xu X, Pasha MA, et al. Cutting edge: notch signaling promotes the plasticity of group-2 innate lymphoid cells. J Immunol. 2017;198(5):1798–1803. doi:10.4049/jimmunol.1601421
136. Kobayashi T, Voisin B, Kim DY, et al. Homeostatic Control of Sebaceous Glands by Innate Lymphoid Cells Regulates Commensal Bacteria Equilibrium. Cell. 2019;176(5):982–997.e916. doi:10.1016/j.cell.2018.12.031
137. To TT, Oparaugo NC, Kheshvadjian AR, et al. Understanding type 3 innate lymphoid cells and crosstalk with the microbiota: a skin connection. Int J mol Sci. 2024;25(4):2021. doi:10.3390/ijms25042021
138. Fujimoto M, Yokoyama M, Kiuchi M, et al. Liver group 2 innate lymphoid cells regulate blood glucose levels through IL-13 signaling and suppression of gluconeogenesis. Nat Commun. 2022;13(1):5408. doi:10.1038/s41467-022-33171-6
139. Ahmed A, Joseph AM, Zhou J, et al. CTLA-4-expressing ILC3s restrain interleukin-23-mediated inflammation. Nature. 2024;630(8018):976–983. doi:10.1038/s41586-024-07537-3
140. Suffiotti M, Carmona SJ, Jandus C, et al. Identification of innate lymphoid cells in single-cell RNA-Seq data. Immunogenetics. 2017;69(7):439–450. doi:10.1007/s00251-017-1002-x
141. Vo DN, Yuan O, Kanaya M, et al. A temporal developmental map separates human NK cells from noncytotoxic ILCs through clonal and single-cell analysis. Blood Adv. 2024;8(11):2933–2951. doi:10.1182/bloodadvances.2023011909
142. Fumagalli V, Venzin V, Di Lucia P, et al. Group 1 ILCs regulate T cell-mediated liver immunopathology by controlling local IL-2 availability. Sci Immunol. 2022;7(68):eabi6112. doi:10.1126/sciimmunol.abi6112
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Primary Cutaneous Anaplastic Large Cell Lymphoma Arising in a Patient with Rhupus Syndrome and Sjogren’s Syndrome
Gao Z, Xu Q, Chen X, Mao D, Zhang J, Jin J
Clinical, Cosmetic and Investigational Dermatology 2022, 15:975-979
Published Date: 30 May 2022
Acthar Gel (RCI): A Narrative Literature Review of Clinical and Economic Evidence
Wan GJ, Niewoehner J, Hayes K
ClinicoEconomics and Outcomes Research 2023, 15:499-512
Published Date: 26 June 2023
Genetically Proxied Autoimmune Diseases and the Risk of Facial Aging
Zhang Z, Li M, Geng Y, Wang W, Wang W, Shao Y
Clinical, Cosmetic and Investigational Dermatology 2024, 17:981-991
Published Date: 3 May 2024
Health Disparities in Rheumatology in the United States
Wright GC, Zueger PM, Copley-Merriman C, Khan S, Costello J, Krumbach A, Reddy P, Tanjinatus O, Wells AF
Open Access Rheumatology: Research and Reviews 2025, 17:1-12
Published Date: 9 January 2025
Role of Gut Microbiota in the Development of Some Autoimmune Diseases
Cui X, Cong Y
Journal of Inflammation Research 2025, 18:4409-4419
Published Date: 25 March 2025