")
Back to Journals » Journal of Inflammation Research » Volume 17
Early-Life Stress Induced by Neonatal Maternal Separation Leads to Intestinal 5-HT Accumulation and Causes Intestinal Dysfunction
Authors Yang D, Bai R , Li C, Sun Y, Jing H, Wang Z, Chen Y, Dong Y
Received 23 July 2024
Accepted for publication 14 November 2024
Published 19 November 2024 Volume 2024:17 Pages 8945—8964
DOI https://doi.org/10.2147/JIR.S488290
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Ding Yang,1 Rulan Bai,1 Chengzhong Li,2 Yan Sun,2 Hongyu Jing,1 Zixu Wang,1 Yaoxing Chen,1 Yulan Dong1
1College of Veterinary Medicine, China Agricultural University, Beijing, People’s Republic of China; 2Department of Horticulture and Landscape Architecture, Jiangsu Agri-Animal Husbandry Vocational College, Taizhou, People’s Republic of China
Correspondence: Yulan Dong, Email [email protected]
Background: The early childhood period is a critical development stage, and experiencing stress during this time may increase the risk of gastrointestinal disorders, including irritable bowel syndrome (IBS). Neonatal maternal separation (NMS) in rodent models has been shown to cause bowel dysfunctions similar to IBS, and 5-HT is considered to be a key regulator regulating intestinal function, but the precise underlying mechanisms remain unclear.
Results: We established a maternal separation stress mouse model to simulate early-life stress, exploring the expression patterns of 5-HT under chronic stress and its mechanisms affecting gut function. We observed a significant increase in 5-HT expression due to NMS, leading to disruptions in intestinal structure and function. However, inhibiting 5-HT reversed these effects, suggesting its potential as a therapeutic target. Furthermore, our research revealed that excess 5-HT in mice with early life stress increased intestinal neural network density and promoted excitatory motor neuron expression. Mechanistically, 5-HT activated the Wnt signaling pathway through the 5-HT4 receptor, promoting neurogenesis within the intestinal nervous system.
Conclusion: These findings shed light on the intricate changes induced by early life stress in the intestines, confirming the regulatory role of 5-HT in the enteric nervous system and providing potential insights for the development of novel therapies for gastrointestinal disorders.
Keywords: 5-HT, IBS, early-life stress, neurogenesis
Introduction
Prolonged exposure to adverse life circumstances such as poverty and inadequate parental support can induce sustained activation of the body’s stress response mechanisms. This enduring activation results in alterations in neural synaptic plasticity,1 subsequently resulting in cognitive impairments, anxiety, and depression.2 Numerous studies on animal models have also shown that early life stress (ELS) induces alterations in the response of animals to environmental stressors. These alterations include changes in cortisol levels and autonomic nervous system responses during laboratory stresses,3 epigenetic changes,4 increased inflammatory activity, and markers of immune dysregulation.5
Furthermore, ELS increases the likelihood of developing functional gastrointestinal disorders (including IBS) later in life.6 IBS is characterized by alterations in bowel habits and heightened sensitivity to intestinal distension,7 and these symptoms are commonly associated with the neural innervation activity of the enteric nervous system (ENS). Previous studies have shown heightened neural connections within the intestinal mucosa and increased sensitivity of afferent pathways in model mice experiencing post-infection and post-inflammatory discomfort,8 suggesting that the enteric nervous system plays a role in the perception of intestinal distress.
The altered metabolism of intestinal serotonin (5-HT) is recognized as a biomarker for IBS.9 5-HT is known to regulate bowel movement and pain perception under normal physiological conditions.10 Recent research has highlighted the role of alterations in 5-HT signaling in gastrointestinal functional disorders, including IBS, chronic constipation, diarrhea, and functional dyspepsia.11 Experimental studies have demonstrated that colonic and intraperitoneal administration of 5-HT can induce intestinal hyperalgesia.12
MIWA et al reported significantly increased mucosal concentrations of 5-HT in IBS patients with constipation than in both healthy individuals and IBS patients with diarrhea.13 Aberrant 5-HT signaling has been associated with gastrointestinal dysmotility, increased visceral organs sensitivity, and abnormalities in intestinal secretions.14 Serotonergic neurons that mature earlier support the development or survival of some late-differentiating enteric neurons, such as dopaminergic neurons.15 Research suggests that the activation of 5-HT4 receptors promotes the proliferation and migration of enteric neurons, thus contributing to the growth and maintenance of the enteric nervous system after birth.16 Enteric glial cells can also generate neurons both in vitro and in vivo through a 5-HT4-dependent mechanism.17 Nonetheless, the specific mechanisms by which serotonin and its receptors influence gastrointestinal motility and function in patients with IBS remain poorly understood. Understanding these mechanisms is crucial for developing targeted therapeutic interventions. Such insights could enable the design of more effective treatments for gastrointestinal disorders related to serotonergic dysregulation, including motility disturbances and visceral hypersensitivity. By elucidating these pathways, we can ultimately provide a stronger foundation for improving treatment outcomes for individuals suffering from stress-induced gastrointestinal dysfunction.
The neonatal maternal separation (NMS) model in rodents is a well-established experimental approach for investigating ELS. It has been shown to induce gastrointestinal dysfunction in rodents during adulthood, including colonic hyperalgesia, increased permeability, and enhanced motility.18 Unlike prior works that broadly correlate ELS with IBS or 5-HT dynamics with psychological disorders, our investigation delves into the specific alterations in intestinal structure, function, and the ENS engendered by 5-HT dysregulation. Our study revealed that ELS significantly elevated 5-HT levels in the gastrointestinal tract, thereby disrupting both intestinal structural and functional homeostasis. Additionally, 5-HT influenced the ENS by increasing the density of the intestinal neural network, increasing the number of excitatory neurons, and stimulating the secretion of related neurotransmitters. These findings reveal the regulatory role of serotonin in the enteric nervous system, which could be a potential mechanism underlying gastrointestinal function disorders and provide valuable insights for studying the pathogenesis of IBS.
Materials and Methods
Animals and Treatments
Pregnant C57BL/6J mice (20–25 g) were obtained from Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). These animals were kept under standardized conditions, including controlled temperature, humidity, and 12-hour light-dark cycles, with ad libitum access to standard chow and tap water. The animal handling protocols followed the criteria for the Care and Use of Laboratory Animals set by China Agricultural University. These procedures were approved by the Institutional Animal Care and Use Committee of China Agricultural University, with permit number AW50204202-2-2 (Beijing, China).
To investigate serotonin expression in mice using the NMS model, animals were divided into control and NMS groups. The NMS mouse model was established as described previously.19 Starting from the 3rd to the 14th day after birth, the pups were removed from their cages daily for a 3-hour period (from 9:00 a.m. to 12:00 a.m.) and then peacefully reunited with their mothers. The control group was raised normally (see Figure 1A for schematic). To maintain consistency, only healthy neonatal pups born to dams with no health abnormalities were included in the study, while pups with visible congenital anomalies or significant health problems were excluded. Additionally, female mice were culled after determining sex at weaning, and only male mice were kept and raised to adulthood for subsequent testing.
 |
Figure 1 ELS induced an increase in 5-HT in the gut. (A) Experimental design diagram of the animal models. (B) Daily body weight. (C) Weekly body weight. (D) Colon and (E) serum 5-HT levels were determined by ELISA (n=6). (F) Immunofluorescence staining of 5-HT in the colon (5-HT, red) and nucleic acid (Dapi, blue) (scale bar=100 μm), and the number of 5-HT-positive cells was quantified. (G) Determination of tryptophan in the colon by UHPLC-MS/MS (n=5). (H) Immunohistochemical staining was performed to detect in colon sections (scale bar=50 μm), and the IOD of CgA-positive cells in the colon was quantified. (I) TPH1 and TPH2 mRNA expression in the colon and muscularis (n=6). (J) SERT and GAPDH protein expression in the colon and muscularis (n=3). Data is presented as the mean ± SEM. *P < 0.05; ***P < 0.001. Abbreviations: Control, control group; NMS, neonatal maternal separation group. |
To investigate the impact of 5-HT on NMS mice, animals were further divided into three groups: control, NMS, and NMS+PCPA. The pups in the NMS+PCPA group were intraperitoneally injected with PCPA (300 mg/kg, C6506, Sigma‒Aldrich, Shanghai, China, dissolved in 0.5% CMC-Na/saline water) at 8:00 a.m. from the 3rd to the 14th day after birth and then separated from their mother. The control and NMS groups received vehicle injections at the same time, with the NMS group treated as before and the control group receiving standard care (see Figure 2A for schematic).
 |
Figure 2 ELS-induced intestinal barrier disruption was associated with 5-HT. (A) Experimental design diagram of the animal models. (B) Colon and (C) serum 5-HT levels were determined by ELISA (n=6). (D) Small intestine length and (E) colon length in mice. (F) H&E staining of the colon (scale bar=100 μm). (G) Claudin-1 immunohistochemical staining of colon sections (scale bar=50 m) and the IOD of claudin-1-positive cells in the colon. (H) The mRNA expression of Claudin-1, Claudin-3, and Occludin in the colon (n=6). (I) The protein expression of Claudin-1, Claudin-3, Occludin, and GAPDH in the colon (n=3). (J) Immunofluorescence staining of F4/80 in the colon (F4/80, red) and nucleic acid staining (Dapi, blue) (scale bar=100 μm) were performed, and the number of F4/80-positive cells was quantified. Data is presented as the mean ± SEM. *P < 0.05; **P < 0.01; ***P < 0.001. Abbreviations: Control, control group; NMS, neonatal maternal separation group; NMS+Pcpa group, neonatal maternal separation + Pcpa intervention group. |
The pups in each group were weaned on the 21st day after birth. In all experiments, the animals were humanely euthanized at 8 weeks of age by inhalation of CO2 to facilitate phenotype analysis.
In vivo Measurements
Fecal Composition
Each mouse was housed individually in cages without bedding, and fecal pellets were collected continuously for one hour and weighed to determine the wet weight. Subsequently, the collected fecal pellets were meticulously dried at 50 °C to determine the dry weight. The dry-wet ratio and fecal water content are then calculated using the wet weight and dry weight measurements described earlier.20
Colonic Motility
The motility of the distal colon was performed by a bead expulsion test according to previous descriptions.21 After anesthetizing the mice with isoflurane (792632, Sigma‒Aldrich, Shanghai, China), a 3 mm diameter glass bead was inserted into the colon 2 cm from the anal edge. The time it took for the mice to expel the glass beads was measured. Each animal underwent the experiment three times with intervals of more than one hour between each trial.
Total Gastrointestinal Transit Time
Total gastrointestinal transit time was measured using nonabsorbable carmine red as described.22 Mice were orally administered 200 μL of a 6% carmine red solution (C6152, Sigma‒Aldrich, Shanghai, China). The duration between oral administration and the initial detection of red-colored feces was documented to determine the total gastrointestinal transit time.
Assessment of Colonic Visceral Sensitivity
Abdominal withdrawal reflex (AWR) tests were conducted to assess colonic visceral sensitivity in mice.19 After the mice were anesthetized with isoflurane (2.5%, 792632, Sigma‒Aldrich, Shanghai, China), an inflated balloon was introduced into the rectum approximately 1 cm from the anus. The measurement electrodes of the electromyography system were attached to the external oblique muscle of the mouse’s abdomen. Once the mice had fully regained consciousness, the balloon was inflated for 10 seconds at a pressure of 50 mmHg, and the electromyographic activity was recorded. The experiment was repeated four times, with 5-minute intervals between each repetition.
Determination of the Concentration of 5-HT
The quantification of 5-HT levels in mouse blood and colon tissue was performed using an ELISA-based method (E-EL-0033c; Laibotairui Tech Co., Ltd., Beijing, China) according to the manufacturer’s instructions according to our published work.23
Assessment of Intestinal Permeability
The assessment of intestinal permeability in mice was conducted utilizing FITC-labeled dextran with a molecular weight of 3000–5000 Da (FD4, Sigma‒Aldrich, Shanghai, China) as described.24 Following an overnight period of fasting, the mice were administered an oral gavage of FITC-labeled dextran (60 mg/100 g of body weight). Serum samples were obtained at 5 hours after oral administration to quantify the fluorescence intensity using a microplate reader (BioTek Co., Ltd., Beijing, China). The fluorescence was obtained with an excitation wavelength of 492 nm, and the emitted light was measured at 525 nm.
Quantitative Detection of Neurotransmitters and Tryptophan
UHPLC-MS/MS has the advantages of high resolution and high sensitivity when used to assess compounds. Here, colon tissue samples were spiked with a 10-fold volume (Vm, μL/mg) of 80% methanol and homogenized by using a Tissuelyser (JX-24, Jingxin, Shanghai, China) with beads at 40 hz for 4 min, after which the mixture was centrifuged at 15000 rcf and 4 °C for 15 min. Then, 40 μL of the supernatant was mixed with 20 μL of sodium carbonate (100 mm). Derivatization was performed at room temperature for 10 min after spiking 20 μL of 2% benzoyl chloride in acetonitrile. After centrifugation at 15000 rcf and 4 °C for 15 min, the supernatant was isometrically mixed with stable isotope-labeled internal standards to quantify the level of amino acids by UHPLC-MS/MS according to a previously described method.25 Nine compounds, including 5-hydroxytryptamine (5-HT), epinephrine (E), norepinephrine (NE), dopamine (DA), γ-aminobutyric acid (GABA), histamine (Hist), asparagine (Asn), acetylcholine (Ach) and tryptophan (Trp), were assessed and their levels were quantified in colon tissue samples.
Histological Staining
The intestinal tissue was washed with PBS, fixed with a 4% paraformaldehyde solution, and subsequently embedded in paraffin according to we described earlier.26 The intestinal sections, with a thickness of 5 μm, were obtained by employing a semiautomatic rotary microtome. Hematoxylin and eosin (H&E) staining was performed on intestinal tissue sections. Immunohistochemistry was performed to determine the presence of Claudin-1 (1:500, ab15098, Abcam, Cambridge, CA, USA) in paraffin-embedded intestinal sections. The sections were incubated overnight at 4 °C. Following the procedure, the sections were subjected to a wash with PBS at a pH of 7.4. Subsequently, the sections were incubated sequentially with biotinylated goat anti-rabbit IgG secondary antibodies (1:300, A0277, Beyotime Co., Ltd., China) at ambient temperature for 2 hours. Subsequently, after the washing step, the samples were incubated sequentially with a 1:300 dilution of HRP-streptavidin (A0303, Beyotime Co., Ltd., China) for 2 hours. An immunological response was observed following treatment with the DAB Kit (ZLI-9017, Zhongshan Jinqiao Biotech Co., Inc., Beijing, China), and the nucleus was counterstained with hematoxylin for 5 minutes. The cytoplasm or nucleus exhibited a yellow‒brown immunoreactive region. The samples were visually examined and imaged using a microscope (BX51, Olympus, Tokyo, Japan). The integrated optical density (IOD) of the positive cells was measured using Image-Pro Plus software (Media, Rockville, MD, USA). Immunofluorescence was performed using specific antibodies against 5-HT (1:300, bs-1126R, Bioss, Beijing, China), CgA (1:300, SC-13090, Santa Cruz, Dallas, TX, USA) and Tuj1 (1:300, ab18207, Abcam, Cambridge, CA, USA) in paraffin-embedded intestinal sections. The staining was carried out overnight at 4 °C. Subsequently, the tissues were decolorized on a shaker using phosphate-buffered saline (PBS). Next, the tissues were subjected to incubation with the appropriate secondary antibody, Goat Anti-Rabbit Alexa fluor 594 (1:300, ab150080, Abcam, Cambridge, CA, USA) or Goat Anti-Mouse Alexa fluor 488 (1:300, ab150113, Abcam, Cambridge, CA, USA), at ambient temperature for 1 hour. The samples were imaged by microscopy (TE 2000S, Nikon Instruments Co., Inc., New York, NY, USA). Using Image-Pro Plus software (Media Cybernetics, Rockville, Maryland, United States), positively stained puncta were quantified.
Immunofluorescence of the Myenteric Plexuses
The colonic plexus whole-mount preparations were obtained following the previously described method,27 with the colon being harvested and then preserved in a 4% formaldehyde solution. The tissues were dissected along the mesenteric boundary, with careful attention to orienting the serosal side in a downward position. The mucosa/submucosa layers and circular muscle were meticulously excised to obtain whole mounts of the longitudinal muscle/myenteric plexus (LMMP). After successful LMMP acquisition, the collected samples were processed for immunohistochemistry analysis. Initially, the samples were subjected to incubation with specific antibodies, namely, mouse anti-HuC/D antibody (1:300, A21271, Invitrogen, Carlsbad, CA, USA), rabbit anti-Tuj1 antibody (1:300, ab18207, Abcam, Cambridge, CA, USA), rabbit anti-S100B antibody (1:500, ab115803, Abcam, Cambridge, CA, USA), rabbit anti-Chat antibody (1:300, A01192-4, Boster, Wuhan, China), and rabbit anti-nNOS antibody (1:500, ab76067, Abcam, Cambridge, CA, USA), for 48 hours at 4 °C. Solution containing the primary antibody was extracted, and the samples were subjected to three washes of 5 minutes each using PBS containing 0.1% Triton X-100. Subsequently, the samples were incubated with the appropriate secondary antibodies, namely, goat anti-rabbit Alexa Fluor 594 (ab150080, Abcam, Cambridge, CA, USA) and goat anti-mouse Alexa Fluor 488 (ab150113, Abcam, Cambridge, CA, USA), for 1 to 2 hours at room temperature. Next, the samples were washed three times for 5 minutes each using PBS. During the process of double labeling, the samples were washed three times with PBS, with each wash lasting for 10 minutes. Subsequently, the second labeling was carried out using an identical methodology. The samples were imaged by microscopy (TE 2000S, Nikon Instruments Co., Inc., New York, NY, USA). The quantification of positively stained puncta was performed using Image-Pro Plus software (Media Cybernetics, Rockville, MD, USA).
NADPH-d Staining
Histochemical analysis was employed to detect the activity of NADPH-d as described,28 and the results revealed the presence of neurons expressing nitric oxide synthase (NOS) in the gastrointestinal tract. The LMMP preparations were washed with PBS, followed by incubation in phosphate buffer (0.1 M, pH 7.4) containing 0.3% Triton X-100, 0.5 mg/mL nitroblue tetrazolium (N6876, Sigma‒Aldrich, Shanghai, China), and 1.0 mg/mL β-NADPH (N7505, Sigma‒Aldrich, Shanghai, China) for 1 h at 37 °C. The progression of the reaction was arrested through the implementation of a technique involving immersion of the sample in PBS.
Cell Culture
To determine the precise mechanism by which 5-HT affects nerve cells, a mouse neuroblastoma N2a cell line was used for in vitro research as previously described.29 N2a cells (ATCC#CCL-131) were cultured in Dulbecco’s modified Eagle’s medium (Vivacell, Shanghai, China) supplemented with 10% FBS, 100 units/mL penicillin, and 0.1 milligrams/mL streptomycin. In this study, cells were initially seeded at a density of 2 × 103 cells/mL in 12-well plates and allowed to incubate overnight. Subsequently, the cells were subjected to daily treatment with 10 μM all-trans retinoic acid (R2625, Sigma‒Aldrich, Shanghai, China) for 3 days. Subsequently, the cells were incubated with 5-HT (1 μM, H9523, Sigma‒Aldrich, Shanghai, China) along with the 5-HT4 receptor antagonist GR113808 (10 nM, HY-103152, MCE, USA) or the Wnt pathway inhibitor IWP-2 (20 μM, HY-13912, MCE, USA) in serum-free media for 2 days. Cultured samples were obtained to determine gene transcript levels and protein levels.
Real-Time Quantitative PCR
Intestinal tissues were obtained from the mice used in the experiment and subjected to homogenization using TRIzol Reagent (CW0580, CoWin Biotech Co., Inc., Beijing, China). Total RNA was then extracted according to the described guidelines.30 The RNA sample, with a concentration 1 μg, was subjected to reverse transcription to generate cDNA using HiScript II Select qRT SuperMix (R312-02, Vazyme, Nanjing, China). The experiment involved the use of a real-time quantitative PCR technique and employed gene-specific primer sets and SYBR Green master mix (Q141-02; Vazyme, Nanjing, China). Changes in fluorescence were observed using a OneStep Plus device manufactured by Applied Biosystems, located in the United States. The primers utilized are listed in Table S1.
Western Blot Analysis
Protein extraction from the intestine and cell lines was performed using RIPA lysis buffer (R0010, Solarbio Life Sciences, Beijing, China) supplemented with protease and phosphatase inhibitors for 30 minutes as described.26 The protein content was measured using a BCA protein quantification kit, followed by separation of the same amount of protein using a 10–15% SDS‒PAGE gel. Subsequently, the separated proteins were electrotransferred onto a polyvinylidene fluoride membrane. The membrane was then blocked for 1.5 hours at room temperature with a solution of 5% skim milk in Tris-buffered saline with Tween 20 (TBST). The membrane was then incubated overnight at 4 °C with the specific antibodies listed in Table S2. The samples were then incubated at room temperature for 2 hours with HRP-conjugated goat anti-mouse IgG or goat anti-rabbit IgG (Table S2). The bands present on the blots were subjected to scanning and subsequent analysis using the ImageJ program (National Institutes of Health in Bethesda, MD, USA). The band intensity of the target were standardized relative to the band intensity of GAPDH.
Microbial Sequencing
The fecal microbiota was subjected to Illumina NovaSeq sequencing at Baimaike in Beijing, China. Fecal DNA samples were amplified by polymerase chain reaction (PCR) with barcode primers located on the V3-V4 region of the 16S rRNA gene. The purification, quantification, and homogenization processes were conducted on the products to create a sequencing library. The library that was created underwent an initial inspection, and the libraries that met the necessary criteria were subsequently sequenced using the Illumina NovaSeq 6000 platform following a previously established process.25
Statistical Analysis
The data analysis was conducted using the GraphPad Prism 8.0 software package. All data were presented as the mean ± SEM. The normality of the data was tested using the Shapiro–Wilk test. Comparisons were made using an unpaired Student’s t-test or a one-way ANOVA followed by Tukey’s multiple-comparison post-hoc test. The data from animal experiments are obtained from at least three biological replicates, whereas the data from cell experiments are derived from three independent experiments to confirm the results. A significance level of P < 0.05 was set as the threshold for statistical significance.
Results
ELS Promotes an Increase in 5-HT Through ECs Proliferation and SERT Inactivation
To examine the influence of 5-HT on gastrointestinal changes caused by ELS, we used a NMS model. Neonatal rodents were separated from their mothers for 3 hours daily from postnatal days 3 to 14, a critical period for neurodevelopment when the brain-gut axis is highly plastic. The animals were then sacrificed at 2 months of age, representing adulthood, to analyze the long-term effects of ELS on gastrointestinal function. (Figure 1A). First, by tracking weight changes in the mice, we found that NMS mice gained weight slowly during separation. However, this process did not continue into adulthood (Figure 1B and C). The slower weight gain is likely due to reduced feeding behavior during the separation period, as maternal separation limits direct access to maternal nutrition during these critical early developmental days. Consistent with previous findings,19 our results show that NMS has a substantial impact on the upregulation of 5-HT expression in the proximal colons of mice (Figure 1D). In addition, we found that 14-day-old mice that had just experienced NMS also exhibited increased colonic 5-HT levels (Figure S1). The serum exhibited comparable upregulation of 5-HT expression (Figure 1E). Immunostaining using a specific antibody against 5-HT confirmed the increase in 5-HT in the colonic epithelium following exposure to NMS (Figure 1F).
To understand the reason for the increase in 5-HT in the colon of NMS mice, we first measured the content of the 5-HT precursor tryptophan in the colon. There was no difference in the amount of tryptophan in the colon between NMS mice and control mice (Figure 1G). Enteroendocrine cells (ECs) are one of the primary sources of 5-HT in the gastrointestinal tract. In this study, ECs were detected by immunofluorescence staining for chromogranin A (CgA) (Figure 1H), which confirmed that there were more ECs in NMS mice than in control mice. In addition, serotonin transporters (SERTs) and tryptophan hydroxylase (TPH) are involved in 5-HT metabolism. The inactivation of SERT is an essential contributor to the increase in 5-HT content in the gut. Considering the different sources of 5-HT (ECs or neurons), the muscularis was isolated separately, and there was no significant difference in the transcription of TPH1 or TPH2 in the colon or muscularis of NMS mice and control mice (Figure 1I). Surprisingly, the transcription and protein expression of SERT in the colon and muscularis of NMS mice were significantly decreased (Figure 1I and J), which may have aggravated the accumulation of 5-HT in the gut of NMS mice.
Excessive 5-HT Accumulation Induced by ELS Alters the Intestinal Barrier
To investigate the potential role of 5-HT in mediating the phenotypic alterations generated by NMS, mice that were subjected to NMS were intraperitoneally administered Pcpa (Figure 2A). Pcpa is an inhibitor of tryptophan hydroxylase, the rate-limiting enzyme in 5-HT synthesis. Pcpa effectively inhibits the expression of both TPH1 and TPH2, reducing 5-HT synthesis in ECs and enteric neurons. Indeed, Pcpa effectively reduced the levels of 5-HT in the colon and serum (Figure 2B and C). Intestinal anatomical examination revealed that NMS did not affect intestinal length (Figure 2D and E). We further investigated the impact of NMS on intestinal structure. The HE staining results showed no significant differences (Figure 2F). However, immunostaining of claudin-1 in the colons of NMS mice was reduced (Figure 2G); this change was reversed by Pcpa treatment. Consistent with the immunostaining results for claudin-1, at both the transcription and protein expression levels, Pcpa inhibited the decreases in claudin-1, claudin-3, and occludin in the colons of NMS mice (Figure 2H and I). Notably, compared to the control group, NMS mice showed a significant increase in the abundance of the surface protein F4/80 on intestinal macrophages (Figure 2J), as well as increased transcription of pro-inflammatory cytokines IL-1β, IL-6, and TNF-α (Figure S2), indicating that NMS induced inflammation in the intestine. Moreover, the reduction in 5-HT alleviated this inflammatory response. From these results, we inferred that the high levels of 5-HT in the intestines of NMS mice, while not causing significant pathological changes to the intestinal barrier, do affect barrier structure to some extent.
Intestinal microbial homeostasis is an important factor that is related to intestinal physiology and function. The composition of the intestinal microbiota is dynamically regulated by 5-HT metabolism. To investigate the effect of excess 5-HT in NMS on gut microbes, we characterized the microbial composition of fecal samples collected from mice using 16S rRNA gene sequencing analyses. The Venn diagram showed that the colonic flora of the three groups of mice had different numbers of OTUs (Figure 3A). However, alpha diversity according to the Shannon and Simpson indices showed no significant difference in species richness among the three groups of mice (Figure 3B and C). Notably, the abundance of Faecalibaculum increased significantly in the gut of NMS mice, and Pcpa treatment restored the abundance of Faecalibaculum to levels similar to those in the control group (Figure 3D). To identify the particular bacterial taxa relevant to the increase in 5-HT concentration induced by NMS, we utilized the linear discriminant analysis (LDA) effect size (LEfSe) approach. A cladogram representative of the structure of the colonic microbiota is shown. This cladogram highlights the dominant bacteria and the notable variations in taxa observed among the three groups (Figure 3E). The findings indicated that the dominant bacteria in the colon of NMS mice was Erysipelotrichaceae. The administration of Pcpa therapy resulted in considerable suppression of this bacterium, even though the predominant bacterial species differed between the control group and the treatment group. A comparative chart illustrating the significant differences between groups revealed that the administration of NMS resulted in an increase in the abundance of Feacalibaculum and Erysipelotrichaceae in the colon, the levels of which returned to control levels after Pcpa treatment (Figure 3F and G). In conclusion, the increase in 5-HT induced by NMS is related to changes in the intestinal barrier and microbial composition.
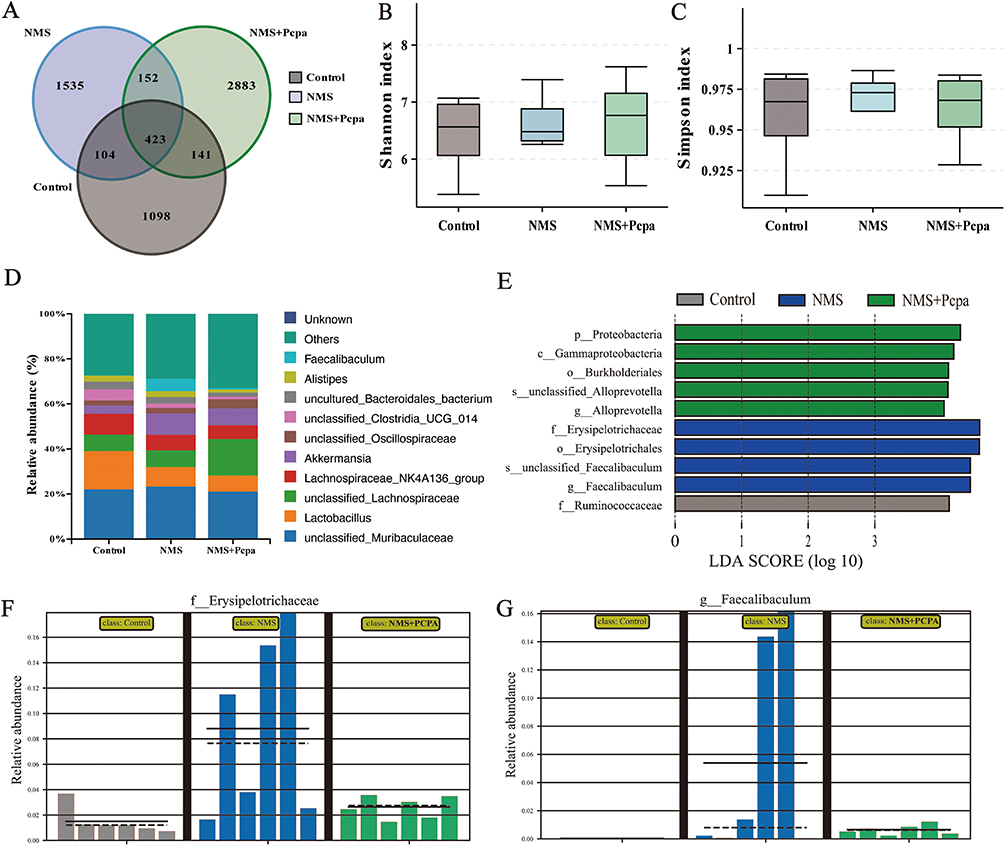 |
Figure 3 ELS altered the gut microbiome. (A) Venn diagram of colonic microbial OTUs. (B) Shannon index of colonic microorganisms. (C) Simpson index of colonic microorganisms. (D) The relative abundance of the top 12 most abundant bacteria at the genus level. (E) Linear discriminant analysis effect size (LEfSe) prediction was used to identify the most differentially abundant bacteria in each group. Relative abundances of Erysipelotrichaceae (F) and Faecalibaculum (G) based on the LefSe results. Abbreviations: Control, control group; NMS, neonatal maternal separation group; NMS+Pcpa group, neonatal maternal separation + Pcpa intervention group. |
Excessive 5-HT Accumulation Induced by ELS Leads to Intestinal Dysfunction
To assess whether the IBS-related symptoms exhibited by NMS mice are associated with 5-HT, we further examined the intestinal function of the mice. We first assessed the integrity of the intestinal barrier by quantifying the leakage of FITC into the circulatory system. Compared with control mice, NMS mice exhibited increased serum FITC levels, which were reduced by Pcpa supplementation, suggesting that the NMS-induced excessive abundance of 5-HT increased intestinal permeability (Figure 4A). Additionally, the water content in excreted feces was evaluated to identify any potential changes in secretion and/or absorption. The experimental group of NMS-exposed mice had greater wet weights of fecal pellets than did the control group (Figure 4B). This observation indicates an increased reaction to stress in NMS mice. Furthermore, there was an increase in the fecal water content in NMS mice (Figure 4C), suggesting potential alterations in water absorption and/or secretion. Similarly, Pcpa treatment improved the stool traits of NMS mice.
 |
Figure 4 Excessive 5-HT accumulation induced by ELS caused intestinal dysfunction. (A) Intestinal permeability (n=5). (B) Fecal pellet wet weight (n=6). (C) Fecal water content (n=6). (D) Whole gut transit time (n=6). (E) Distal colonic motility (n=6). (F) Assessment of the visceral pain threshold in mice (n=6). Data is presented as the mean ± SEM. *P < 0.05; **P < 0.01; ***P < 0.001. Abbreviations: Control, control group; NMS, neonatal maternal separation group; NMS+Pcpa group, neonatal maternal separation + Pcpa intervention group. |
Gastrointestinal motility was subsequently assessed by employing three distinct methodologies. We found that gastrointestinal motility was accelerated in NMS mice, as indicated by a reduction in whole gut transit time (Figure 4D) and a decrease in colon expulsion time (Figure 4E). Pcpa reversed the acceleration of intestinal motility in NMS mice. Finally, we examined visceral hypersensitivity in the gut. Compared with control mice, NMS animals exhibited heightened sensitivity to visceral discomfort during the colorectal distention test, as evidenced by their reduced intraluminal pressure (Figure 4F). The mice that received an intraperitoneal injection of Pcpa exhibited visceral pain thresholds similar to those of the control group (Figure 4F), suggesting that 5-HT may contribute to visceral hypersensitivity. In summary, NMS-induced increases in the abundance of 5-HT altered intestinal function in mice.
Excessive 5-HT Abundance Causes Neuroanatomical Changes in the Enteric Nervous System
Next, we sought to explore the specific mechanisms by which 5-HT interferes with intestinal motility. The signaling cascade of 5-HT requires interactions with 5-HT receptors. Therefore, we assessed the transcription levels of specific 5-HT receptors in the colons of NMS mice. The gut expresses several 5-HT receptors, among which 5-HT4 receptors are highly expressed in the colon of NMS mice (Figure 5A and B). Multiple studies have shown that the 5-HT4 receptor mediates neuroprotection and neurogenesis in the gut.16 We hypothesize that 5-HT-related dysfunction in NMS may be related to the enteric nervous system. This hypothesis was confirmed by the increased transcription levels of genes associated with gut midgut neurodevelopment (Tubb3, Sox2, Nestin, Olig2, and Gfap) in NMS mice (Figure 5C). The transcription levels of Gfap, which is a glial cell marker, were also elevated (Figure 5C). Pcpa inhibited these alterations, indicating that 5-HT may influence neurogenesis in the enteric nervous system of NMS mice. Importantly, the protein levels of Tuj1 and HUC/D were increased in the gut of NMS mice (Figure 5D). Similarly, there was an increase in the number of myenteric neurons and innervation of the LMMP in NMS animals. This change was determined using immunohistochemistry quantification of the pan-neuronal marker HUC/D and the neurite marker Tuj1 (Figure 5E–G). In addition, NMS mice showed increased colonic epithelial innervation (Figure 5H and I) and glial network density (as revealed by immunostaining for the glia-specific marker S100β; Figure 5J and K). These phenotypes were restored after Pcpa treatment. This finding suggested that excess 5-HT in NMS increases the density of the intestinal neural network.
 |
Figure 5 5-HT promoted neurogenesis in the enteric nervous system. (A) The mRNA expression of 5-HT3, 5-HT4, and 5-HT7 in the colon (n=6). (B) The protein expression of 5-HT4 and GAPDH in the colon (n=3). (C) The mRNA expression of Tubb3, Sox2, Nestin, Olig2, and Gfap in the muscularis (n=6). (D) The protein expression of Tuj1, HUC/D, and GAPDH in the muscularis (n=3). (E) Immunofluorescence staining of HUC/D (green) and Tuj1 (red) in the LMMP (scale bar=100μm), and the number of HUC/D (F) and Tuj1 (G) positive cells was quantified. (H) Immunofluorescence staining of Tuj1 in the colon (Tuj1, red) and nucleic acid (Dapi, blue) (scale bar=100 μm), and (I) the number of Tuj1-positive cells was quantified. (J) Immunofluorescence staining of S100β in LMMP (S100β, red) and nucleic acid (Dapi, blue) (scale bar=100 μm), and (K) the number of S100β-positive cells was quantified. Data is presented as the mean ± SEM. *P < 0.05; **P < 0.01; ***P < 0.001. Abbreviations: Control, control group; NMS, neonatal maternal separation group; NMS+Pcpa group, neonatal maternal separation + Pcpa intervention group. |
The NMS-Induced Increase in 5-HT Promotes the Expression of Excitatory Neurons in the ENS
The ENS contains a variety of neurotransmitters, and neurons that release different neurotransmitters are classified as excitatory and inhibitory motor neurons according to their functions. The intestines’ excitatory and inhibitory motor neurons coordinate and constrain each other, creating a complex and delicate balance between them. First, we used UHPLC-MS/MS to measure the levels of neurotransmitters in the colon (Figure 6A–H). Consistent with previous findings, NMS significantly increased the levels of 5-HT in the mouse colon (Figure 6A). The levels of excitatory neurotransmitters (Ach and asparagine) were significantly elevated in NMS mice (Figure 6B and C). On the other hand, the expression of the inhibitory neurotransmitter GABA was downregulated considerably (Figure 6D). Dopamine levels also exhibited a decreasing trend (Figure 6E), while norepinephrine (Figure 6F), epinephrine (Figure 6G), and histamine (Figure 6H) levels remained unaffected by NMS. We further explored the changes in different neuronal types in the ENS. Immunofluorescence staining of LMMPs revealed that the intestinal acetylcholinergic neurons in NMS mice increased significantly, while Pcpa decreased the number of Chat-positive cells (Figure 6I and J). In contrast, the inhibitory neurons represented by nNOS was reduced in NMS mice (Figure 6K and L). NADPH staining also confirmed these results (Figure 6M and N). In conclusion, these results suggest that the abnormal bowel motor function induced by NMS may be related to an increase in excitatory neurons and a decrease in inhibitory neurons mediated by 5-HT.
 |
Figure 6 ELS altered the types of neurons in the ENS. (A–Η) of the levels of neurotransmitters in the colon was determined by UHPLC-MS/MS (n=6). (I) Immunofluorescence staining of Chat in LMMP (Chat, red) and nucleic acid (Dapi, blue) (scale bar=100 μm), and (J) the number of Chat-positive cells was quantified. (K) Immunofluorescence staining of HUC/D (green) and nNOS (red) in the LMMP (scale bar= 100 μm), and (L) the number of HUC/D- and nNOS-positive cells was quantified. (M) LMMP was subjected to NADPH-d histochemistry (scale bar= 100 μm), and (N) the number of nitrergic neurons was quantified. Data is presented as the mean ± SEM. *P < 0.05; **P < 0.01; ***P < 0.001. Abbreviations: Control, control group; NMS, neonatal maternal separation group; NMS+Pcpa group, neonatal maternal separation + Pcpa intervention group. |
5-HT Activates the Wnt Pathway Through the 5-HT4 Receptor and Promotes Neurogenesis
The effect of 5-HT on the intestinal neural network in NMS mice was investigated. We wanted to explore the specific mechanisms by which 5-HT promotes neurogenesis. In the gut, 5-HT can activate the Wnt signaling pathway and promote the proliferation of colon stem cells.31 We hypothesize that 5-HT promotes neurogenesis through the Wnt pathway. Hence, we examined the expression of Wnt signaling-related molecules in the colonic muscular layer of NMS mice. Compared with that in the control group, the Wnt signaling pathway was activated in the muscle layer of NMS mice, which showed an increase in the level of β-catenin and a decrease in the levels of its degradation complexes APC, Axin1, and GSK-3β (Figure 7A and 7B). These levels recovered to levels comparable to that of the control group after Pcpa treatment. Our previous results showed that the level of the 5-HT4 receptor was significantly elevated. Is 5-HT activation of the Wnt pathway mediated by the 5-HT4 receptor? To answer this question, we studied the effect of the 5-HT4 receptor on the Wnt pathway using mouse neuron cell lines. N2a neuroblastoma cells were incubated with 5-HT to verify the role of the 5-HT4 receptor. Compared with those in the control group, 5-HT promoted the activation of β-catenin (Figure 7C). However, the 5-HT4 receptor antagonist GR113808 inhibited 5-HT-induced activation of β-catenin (Figure 7C), thus demonstrating that in N2a cells, the 5-HT4 receptor mediates activation of the Wnt pathway.
 |
Figure 7 5-HT4 mediates 5-HT activation of the Wnt signaling pathway to promote neurogenesis. (A and B) The protein expression levels of β-catenin, APC, Axin1, GSK-3β, and GAPDH in the muscularis muscle were determined by Western blotting (n=3). (C) Western blotting was used to determine the protein expression of β-catenin, and GAPDH in N2a cells (n=3). (D) The expression of Tubb3, Sox2, and Nestin in N2a cells was measured by real-time qPCR (n=6). (E–H) The protein expression of β-catenin, HUC/D, Tuj1, and GAPDH in N2a cells was examined by Western blotting, and the relative protein levels were normalized to those of GAPDH (n=3). Data is presented as the mean ± SEM. *P < 0.05; **P < 0.01. Abbreviations: Control, control group; NMS, neonatal maternal separation group; NMS+Pcpa group, neonatal maternal separation + Pcpa intervention group. |
To further validate the impact of 5-HT and the Wnt pathway on neural cells, we assessed the expression of neural markers in N2a cells. The results revealed that GR113808 and the Wnt inhibitor IWP-2 significantly suppressed the increase in the transcription of neurogenesis-related factors induced by 5-HT in N2a cells (Figure 7D). Compared to cells treated with 5-HT, cells treated with GR113808 and IWP-2 exhibited decreased protein levels of Tuj1 and HUC/D (Figure 7E–H). In conclusion, 5-HT activates the Wnt pathway through the 5-HT4 receptor and promotes neurogenesis.
Discussion
IBS is a significant clinical issue, as its symptoms are believed to be associated with changes in gastrointestinal motility and/or increased sensitivity in the visceral region;32 furthermore, there could be an excessive release of neuroendocrine transmitters either in the peripheral or central regions in patients with IBS.33 Additionally, combined social- or environment-driven psychological factors, including depression and anxiety, could shape the manifestation of symptoms in individuals with IBS.34 Multiple studies have suggested alterations in serotonin signaling in IBS-induced gastrointestinal dysfunction.35 In the present study, we provided experimental evidence that the neurotransmitter 5-HT drives changes in intestinal function, primarily through the promotion of neurogenesis in the intestinal nervous system. Our study revealed that the use of mother-child separation stress as a standard experimental model of IBS induced an increase in the secretion of 5-HT in the intestine, resulting in increased innervation of the intestinal nervous system, accompanied by an increase in excitatory neurotransmitters and related neurons, ultimately leading to changes in intestinal function.
Several studies have reported abnormal metabolism of 5-HT in the gut of patients with IBS. Previous studies have documented changes in ECs in individuals with “post dysenteric” IBS characterized by recurring stomach pain and diarrhea.36 The presence of ECs hyperplasia and the dysregulation of 5-hydroxytryptamine secretion by these cells has also been reported in the colonic tissues of IBS patients.37 The results of this study indicate that there is a potential link between modified ECs and the manifestation of symptoms in individuals with diarrhea-predominant irritable bowel syndrome (D-IBS). In our study, NMS mice showed elevated levels of 5-HT in the gut and serum, and the number of ECs in the colon was significantly greater in the NMS group than in the control group. This finding is consistent with previous research. Furthermore, there was a notable reduction in the expression of SERT in biopsy samples obtained from individuals diagnosed with IBS compared to those from healthy individuals. The infusion of fecal supernatant from patients with IBS-D into the colon of mice led to an elevation in 5-HT levels and a reduction in the expression of SERT in the colon, as reported in a previous study.38 Similarly, our results revealed SERT inactivation in the colon and muscle of NMS mice. Notably, the level of the rate-limiting synthase TPH1/TPH2 of 5-HT did not change significantly. Our results further revealed the metabolic pathway of 5-HT and revealed that the increase in 5-HT may be caused by an increase in the number of ECs and SERT inactivation.
Currently, most ELS studies have focused on brain neurodevelopment, such as emotion and cognition.39 However, further investigation is required to fully understand the effects of ELS on the structure of the intestines. The intestinal barrier serves as a crucial link between the intestinal microbiota and the immune system of the intestines; thus, this barrier plays a vital role in the preservation of mucosal homeostasis.40 The presence of compromised intestinal barrier function is frequently observed in a range of gastrointestinal illnesses, such as IBS.41 The primary mechanism responsible for maintaining the integrity of the intestinal barrier is tight junctions between epithelial cells. These tight junctions play a crucial role in preventing the entry of hazardous substances such as infectious agents and endotoxins into the bloodstream through the intestinal mucosa.42 A decrease in the expression of tight junction proteins has been associated with increased colon permeability in individuals with IBS,43 which may be a potential cause of visceral sensitivity and abdominal pain in IBS patients. In addition, 5-HT in the gut promotes inflammation and can lead to inflammatory bowel disease.44 Our findings indicate that NMS induces the activation of colonic macrophages and an increase in the transcription of inflammatory cytokines, suggesting the occurrence of colonic inflammation. The expression of tight-linking proteins was also downregulated in the colon of NMS mice. These phenomena were reversed after 5-HT was inhibited. Gut microbes are also essential parts of the intestinal barrier, and the regulatory effect of 5-HT on gut microbes has been gradually revealed in gut-related diseases. According to our results, 5-HT significantly alters the composition of the intestinal flora, and the abundances of Faecalibaculum and Erysipelotrichaceae are influenced by the level of 5-HT, suggesting that the impact of 5-HT on intestinal microbes may be a potential cause of IBS. However, whether specific microbial changes are related to intestinal dysfunction caused by NMS still needs to be further explored. In summary, we demonstrated that 5-HT changed the gut barrier structure in an IBS rodent model.
Despite the three types of IBS, constipation-dominant irritable bowel syndrome (C-IBS), D-IBS, and alternating types, bowel function in patients with any type of IBS is related to bowel frequency and fecal concentration.32 The defecation frequency depends on complex codependent factors, including rectal filling, rectal sensation, socialization, and motivational behaviors.45 The physiological processes of colonic transit and subsequent water absorption in the colon play a crucial role in determining the consistency of stool.46 Our results revealed the downregulation of colonic tight junction protein expression in NMS mice, which is directly related to the intestinal permeability of mice. Consistent with these findings, the intestinal permeability and fecal water content of NMS mice were increased and regulated by 5-HT. This result supports our hypothesis that the regulatory effect of 5-HT on the intestinal barrier in IBS models may be a potential factor underlying the altered bowel habits of IBS patients. Notably, the role of aquaporins (AQPs), particularly AQP3 and AQP8, in regulating water transport across intestinal epithelial cells may also contribute to the observed increase in fecal water content. Aquaporins are crucial for maintaining water homeostasis in the gastrointestinal tract, and imbalances in their expression can affect fluid movement, leading to changes in fecal properties.47 Previous studies have found that maternal separation can alter AQP expression.48 Although we did not specifically measure AQP expression in this study, future research could focus on evaluating the effects of NMS and intestinal 5-HT on AQP3 and AQP8 levels to better understand their relationship with the increased fecal water content observed. This may provide deeper insights into the mechanisms of stress-induced intestinal dysfunction. In addition, we observed accelerated intestinal transport and increased visceral sensitivity in NMS mice, which is consistent with previous reports. These effects were reversed by treatment with 5-HT inhibitors, again confirming the role of 5-HT in regulating bowel movement and pain perception in IBS patients.
Notably, 5-HT also affects the intestinal transport speed of NMS mice. Most of our studies on IBS-associated phenotypes were performed on the colon, and this result suggests that 5-HT regulation of IBS may involve the entire intestine, including the small intestine and colon. This idea is worthy of further study. Intestinal motility and visceral hypersensitivity are intricately linked to the innervation of the enteric nervous system.49 5-HT regulates neurogenesis and postinjury repair of the intestinal nervous system through its receptors.16 Nevertheless, there is limited knowledge about the role of the nervous system in ELS-induced intestinal dysfunction. In our research, we found that NMS increased the expression of 5-HT4 receptors and genes associated with neurogenesis in the colon of mice. 5-HT signaling mediated an increase in neural network density and colonic epithelial innervation in the colon of NMS mice. Moreover, we observed an increase in excitatory motor neurons and a decrease in inhibitory neurons in the ENS. To this end, increased intestinal serotonin in NMS mice may lead to remodeling of the intestinal nervous system.
Our study provides valuable insights into the intricate relationships among 5-HT-5-HT4 receptor signaling, the Wnt pathway, and neurogenesis in the ENS. The significance of the 5-HT4 receptor in addressing conditions such as IBS or even neuroprotection has been well recognized in the field. Notably, 5-HT4 receptor stimulation has been associated with augmented neurogenesis within the CNS.50 Moreover, emerging evidence suggests that 5-HT4 receptors play a crucial role in regulating neurogenesis and neuronal survival during the postnatal developmental stage. This notion is reinforced by the absence of the expected increase in enteric neurons during early bowel maturation in mice lacking 5-HT4 receptors, where conspicuous autophagy was observed within the ENS,16 suggesting that 5-HT4 receptors play a vital role in the maintenance of ENS integrity.
Our in vitro experiments using neuronal N2a cells provided further clarity regarding the role of the 5-HT4 receptor in Wnt signaling. Incubation with 5-HT led to increased expression of the 5-HT4 receptor, and the 5-HT4 receptor inhibitor GR113808 confirmed the direct influence of 5-HT4 on Wnt signaling. These findings collectively confirm the role of 5-HT4-Wnt signaling in facilitating the neurogenesis of N2a cells.
Excessive proliferation of intestinal stem cells (ISCs) resulting from overactivation of the Wnt signaling pathway by 5-HT is a crucial mechanism in colorectal cancer.19 In this study, we unveiled an additional facet of 5-HT and Wnt signaling in the context of gastrointestinal health. Activation of the Wnt signal by 5-HT increases the density of the enteric nervous network. These insights, in conjunction with the literature, highlight the potential negative implications of prolonged exposure to elevated 5-HT levels within the gut. Such exposure may compromise gut health and increase susceptibility to gastrointestinal conditions, including IBS, inflammatory bowel disease (IBD), and even colorectal cancer.19
Our study revealed ELS-mediated remodeling of the intestinal nervous system through the 5-HT/5-HT4 receptor-Wnt pathway and revealed the potential risks of disrupting intestinal homeostasis through excessive 5-HT activity. Finally, our investigation provides a comprehensive understanding of how ELS orchestrates alterations in the enteric nervous system through the activation of the Wnt pathway, which is facilitated by 5-HT/5-HT4 receptor signaling. This dysregulation increases susceptibility to intestinal dysfunction, thus emphasizing the importance of targeted interventions.
Conclusion
In conclusion, our research revealed that elevated 5-HT levels in the gut resulting from ELS contribute to intestinal dysfunction in mice. 5-HT, by activating the 5-HT4 receptor, triggers the Wnt signaling pathway, thereby fostering neurogenesis within the intestinal nervous system and augmenting excitatory neurons (Figure 8). This mechanism may underlie alterations in intestinal motility.
 |
Figure 8 Overview of the role of 5-HT signaling in ELS-induced intestinal dysfunction. Elevated serotonin levels in the gut due to early stress contribute to intestinal dysfunction in mice. These effects may occur through activation of the 5-HT4 receptor, stimulation of the Wnt pathway, the promotion of neurogenesis, an increase in excitatory neurons, and subsequent impacts on intestinal motility. |
Abbreviations
5-HT, Serotonin, 5-hydroxytryptamine; Ach, Acetylcholine; Asn, Asparagine; AWR, Abdominal withdrawal reflex; C-IBS, constipation-dominant irritable bowel syndrome; D-IBS, diarrhea-predominant irritable bowel syndrome; DA, dopamine; E, Epinephrine; EC, enteroendocrine cell; ENS, enteric nervous system; GABA, γ-aminobutyric acid; Hist, histamine; HPA, hypothalamic-pituitary-adrenal; IBD, inflammatory bowel disease; IBS, irritable bowel syndrome; ISC, intestinal stem cells; LMMP, longitudinal muscle/myenteric plexus; NE, norepinephrine; NMS, neonatal maternal separation; NOS, nitric oxide synthase; SERT, serotonin transporters; Trp, tryptophan; TPH, tryptophan hydroxylase.
Data Sharing Statement
The data will be available on reasonable request.
Ethics Approval and Consent to Participate
The animal study was approved by the Institutional Animal Care and Use Committee of China Agricultural University and performed according the criteria for the Care and Use of Laboratory Animals set by China Agricultural University.
Acknowledgments
We sincerely thank the anatomical tissue culture research team of China Agricultural University School of Veterinary Medicine. The Figure 8 was created with BioRender.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The current research was supported by the Natural Science Foundation of Beijing Municipality (6212018), the Forestry Science and Technology programs of Jiangsu Province (LYKJ[2020]29), the Modern Agriculture (Flower) Industrial Technology System of Jiangsu Province (JATS[2023]355), and the Chinese Traditional Medicine Germplasm Resources Comprehensive Utilization Engineering Research Center of Taizhou City (TZDRC[2023]133).
Disclosure
The authors declare that they have no competing interests.
References
1. Ganzel BL, Morris PA, Wethington E. Allostasis and the human brain: integrating models of stress from the social and life sciences. Psychol Rev. 2010;117(1):134–174. doi:10.1037/a0017773
2. Sousa C, Mason WA, Herrenkohl TI, Prince D, Herrenkohl RC, Russo MJ. Direct and indirect effects of child abuse and environmental stress: a lifecourse perspective on adversity and depressive symptoms. Am J Orthopsychiatry. 2018;88(2):180–188. doi:10.1037/ort0000283
3. Ansell EB, Rando K, Tuit K, Guarnaccia J, Sinha R. Cumulative adversity and smaller gray matter volume in medial prefrontal, anterior cingulate, and insula regions. Biol Psychiatry. 2012;72(1):57–64. doi:10.1016/j.biopsych.2011.11.022
4. Turecki G, Meaney MJ. Effects of the social environment and stress on glucocorticoid receptor gene methylation: a systematic review. Biol Psychiatry. 2016;79(2):87–96. doi:10.1016/j.biopsych.2014.11.022
5. O’Donovan A, Neylan TC, Metzler T, Cohen BE. Lifetime exposure to traumatic psychological stress is associated with elevated inflammation in the heart and soul study. Brain Behav Immun. 2012;26(4):642–649. doi:10.1016/j.bbi.2012.02.003
6. Agostini A, Rizzello F, Ravegnani G, et al. Adult attachment and early parental experiences in patients with Crohn’s disease. Psychosomatics. 2010;51(3):208–215. doi:10.1176/appi.psy.51.3.208
7. Thompson W, Longstreth G, Drossman D, Heaton K, Irvine E, Muller-Lissner S. Functional bowel disorders and functional abdominal pain. Gut. 1999;45(Suppl 2):II43–II47. doi:10.1136/gut.45.2008.ii43
8. Hsu LT, Hung KY, Wu HW, et al. Gut-derived cholecystokinin contributes to visceral hypersensitivity via nerve growth factor-dependent neurite outgrowth. J Gastroenterol Hepatol. 2016;31(9):1594–1603. doi:10.1111/jgh.13296
9. Brown PM, Drossman DA, Wood AJJ, et al. The tryptophan hydroxylase inhibitor LX1031 shows clinical benefit in patients with nonconstipating irritable bowel syndrome. Gastroenterology. 2011;141(2):507–516. doi:10.1053/j.gastro.2011.05.005
10. Gros M, Gros B, Mesonero JE, Latorre E. Neurotransmitter dysfunction in irritable bowel syndrome: emerging approaches for management. J Clin Med. 2021;10(15):3429. doi:10.3390/jcm10153429
11. Holtmann G, Shah A, Morrison M. Pathophysiology of functional gastrointestinal disorders: a holistic overview. Dig Dis. 2017;35(Suppl 1):5–13. doi:10.1159/000485409
12. Cenac N, Altier C, Motta JP, et al. Potentiation of TRPV4 signalling by histamine and serotonin: an important mechanism for visceral hypersensitivity. Gut. 2010;59(4):481–488. doi:10.1136/gut.2009.192567
13. Miwa J, Echizen H, Matsueda K, Umeda N. Patients with constipation-predominant irritable bowel syndrome (IBS) may have elevated serotonin concentrations in colonic mucosa as compared with diarrhea-predominant patients and subjects with normal bowel habits. Digestion. 2001;63(3):188–194. doi:10.1159/000051888
14. Lu CL, Hsieh JC, Dun NJ, et al. Estrogen rapidly modulates 5-hydroxytrytophan-induced visceral hypersensitivity via GPR30 in rats. Gastroenterology. 2009;137(3):1040–1050. doi:10.1053/j.gastro.2009.03.047
15. Li Z, Chalazonitis A, Huang Y, et al. Essential roles of enteric neuronal serotonin in gastrointestinal motility and the development/survival of enteric dopaminergic neurons. J Neurosci. 2011;31(24):8998–9009. doi:10.1523/JNEUROSCI.6684-10.2011
16. Liu MT, Kuan YH, Wang J, Hen R, Gershon MD. 5-HT4 receptor-mediated neuroprotection and neurogenesis in the enteric nervous system of adult mice. J Neurosci. 2009;29(31):9683–9699. doi:10.1523/JNEUROSCI.1145-09.2009
17. Belkind-Gerson J, Hotta R, Nagy N, et al. Colitis induces enteric neurogenesis through a 5-HT4-dependent mechanism. Inflamm Bowel Dis. 2015;21(4):870–878. doi:10.1097/MIB.0000000000000326
18. Gareau MG, Jury J, Yang PC, MacQueen G, Perdue MH. Neonatal maternal separation causes colonic dysfunction in rat pups including impaired host resistance. Pediatr Res. 2006;59(1):83–88. doi:10.1203/01.pdr.0000190577.62426.45
19. Wong HLX, yan QH, Tsang SW, et al. Early life stress disrupts intestinal homeostasis via NGF-TrkA signaling. Nat Commun. 2019;10:1745. doi:10.1038/s41467-019-09744-3
20. Vicentini FA, Keenan CM, Wallace LE, et al. Intestinal microbiota shapes gut physiology and regulates enteric neurons and glia. Microbiome. 2021;9:210. doi:10.1186/s40168-021-01165-z
21. Li ZS, Schmauss C, Cuenca A, Ratcliffe E, Gershon MD. Physiological modulation of intestinal motility by enteric dopaminergic neurons and the D2 receptor: analysis of dopamine receptor expression, location, development, and function in wild-type and knock-out mice. J Neurosci. 2006;26(10):2798–2807. doi:10.1523/JNEUROSCI.4720-05.2006
22. Kimball ES, Palmer JM, D’Andrea MR, Hornby PJ, Wade PR. Acute colitis induction by oil of mustard results in later development of an IBS-like accelerated upper GI transit in mice. Am J Physiol Gastrointest Liver Physiol. 2005;288(6):G1266–1273. doi:10.1152/ajpgi.00444.2004
23. Ma J, Wang R, Chen Y, Wang Z, Dong Y. 5-HT attenuates chronic stress-induced cognitive impairment in mice through intestinal flora disruption. J Neuroinflammation. 2023;20:23. doi:10.1186/s12974-023-02693-1
24. Dong L, Xie J, Wang Y, et al. Mannose ameliorates experimental colitis by protecting intestinal barrier integrity. Nat Commun. 2022;13:4804. doi:10.1038/s41467-022-32505-8
25. Guo Y, Zhu X, Zeng M, et al. A diet high in sugar and fat influences neurotransmitter metabolism and then affects brain function by altering the gut microbiota. Transl Psychiatry. 2021;11(1):328. doi:10.1038/s41398-021-01443-2
26. Yu J, Liu T, Guo Q, Wang Z, Chen Y, Dong Y. Disruption of the intestinal mucosal barrier induced by high fructose and restraint stress is regulated by the intestinal microbiota and microbiota metabolites. Microbiol Spectr. 2023;11(2):e0469822. doi:10.1128/spectrum.04698-22
27. Yarandi SS, Kulkarni S, Saha M, Sylvia KE, Sears CL, Pasricha PJ. Intestinal bacteria maintain adult enteric nervous system and nitrergic neurons via toll-like receptor 2-induced neurogenesis in mice. Gastroenterology. 2020;159(1):200–213.e8. doi:10.1053/j.gastro.2020.03.050
28. Sun T, Li D, Hu S, et al. Aging-dependent decrease in the numbers of enteric neurons, interstitial cells of Cajal and expression of connexin43 in various regions of gastrointestinal tract. Aging. 2018;10(12):3851–3865. doi:10.18632/aging.101677
29. Gao W, Chen R, Xie N, Tang D, Zhou B, Wang D. Duloxetine-induced neural cell death and promoted neurite outgrowth in N2a cells. Neurotox Res. 2020;38(4):859–870. doi:10.1007/s12640-020-00216-x
30. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25(4):402–408. doi:10.1006/meth.2001.1262
31. Zhu P, Lu T, Chen Z, et al. 5-hydroxytryptamine produced by enteric serotonergic neurons initiates colorectal cancer stem cell self-renewal and tumorigenesis. Neuron. 2022:S0896627322003695. doi:10.1016/j.neuron.2022.04.024
32. Chey WD, Kurlander J, Eswaran S. Irritable bowel syndrome: a clinical review. JAMA. 2015;313(9):949–958. doi:10.1001/jama.2015.0954
33. Chen M, Ruan G, Chen L, et al. Neurotransmitter and intestinal interactions: focus on the microbiota-gut-brain axis in irritable bowel syndrome. Front Endocrinol. 2022;13:817100. doi:10.3389/fendo.2022.817100
34. Shiha MG, Aziz I. Review article: physical and psychological comorbidities associated with irritable bowel syndrome. Aliment Pharmacol Ther. 2021;54(Suppl 1):S12–S23. doi:10.1111/apt.16589
35. Dunlop SP, Coleman NS, Blackshaw E, et al. Abnormalities of 5-hydroxytryptamine metabolism in irritable bowel syndrome. Clin Gastroenterol Hepatol. 2005;3(4):349–357. doi:10.1016/s1542-3565(04)00726-8
36. Spiller RC, Jenkins D, Thornley JP, et al. Increased rectal mucosal enteroendocrine cells, T lymphocytes, and increased gut permeability following acute Campylobacter enteritis and in post-dysenteric irritable bowel syndrome. Gut. 2000;47(6):804–811. doi:10.1136/gut.47.6.804
37. Sikander A, Rana SV, Prasad KK. Role of serotonin in gastrointestinal motility and irritable bowel syndrome. Clin Chim Acta. 2009;403(1–2):47–55. doi:10.1016/j.cca.2009.01.028
38. Gao J, Xiong T, Grabauskas G, Owyang C. Mucosal serotonin reuptake transporter expression in irritable bowel syndrome is modulated by gut microbiota via mast cell-prostaglandin E2. Gastroenterology. 2022;162(7):1962–1974.e6. doi:10.1053/j.gastro.2022.02.016
39. Allen L, Dwivedi Y. MicroRNA mediators of early life stress vulnerability to depression and suicidal behavior. Mol Psychiatry. 2020;25(2):308–320. doi:10.1038/s41380-019-0597-8
40. Rescigno M. The intestinal epithelial barrier in the control of homeostasis and immunity. Trends Immunol. 2011;32(6):256–264. doi:10.1016/j.it.2011.04.003
41. Hanning N, Edwinson AL, Ceuleers H, et al. Intestinal barrier dysfunction in irritable bowel syndrome: a systematic review. Therap Adv Gastroenterol. 2021;14:1756284821993586. doi:10.1177/1756284821993586
42. Kuo WT, Odenwald MA, Turner JR, Zuo L. Tight junction proteins occludin and ZO-1 as regulators of epithelial proliferation and survival. Ann NY Acad Sci. 2022;1514(1):21–33. doi:10.1111/nyas.14798
43. Annaházi A, Ferrier L, Bézirard V, et al. Luminal cysteine-proteases degrade colonic tight junction structure and are responsible for abdominal pain in constipation-predominant IBS. Am J Gastroenterol. 2013;108(8):1322–1331. doi:10.1038/ajg.2013.152
44. Haq S, Wang H, Grondin J, et al. Disruption of autophagy by increased 5-HT alters gut microbiota and enhances susceptibility to experimental colitis and Crohn’s disease. Sci Adv. 2021;7(45):eabi6442. doi:10.1126/sciadv.abi6442
45. Azpiroz F, Enck P, Whitehead WE. Anorectal functional testing: review of collective experience. Am J Gastroenterol. 2002;97(2):232–240. doi:10.1111/j.1572-0241.2002.05450.x
46. Crowell MD. Role of serotonin in the pathophysiology of the irritable bowel syndrome. Br J Pharmacol. 2004;141(8):1285–1293. doi:10.1038/sj.bjp.0705762
47. Ikarashi N, Kon R, Sugiyama K. Aquaporins in the colon as a new therapeutic target in diarrhea and constipation. Int J Mol Sci. 2016;17(7):1172. doi:10.3390/ijms17071172
48. Chen J, Xu XW, Kang JX, Zhao BC, Xu YR, Li JL. Metasilicate-based alkaline mineral water confers diarrhea resistance in maternally separated piglets via the microbiota-gut interaction. Pharmacol Res. 2023;187:106580. doi:10.1016/j.phrs.2022.106580
49. Cibert-Goton V, Lam C, Lingaya M, et al. Pain severity correlates with biopsy-mediated colonic afferent activation but not psychological scores in patients with IBS-D. Clin Transl Gastroenterol. 2021;12(2):e00313. doi:10.14309/ctg.0000000000000313
50. Lucas G, Rymar VV, Du J, et al. Serotonin(4) (5-HT(4)) receptor agonists are putative antidepressants with a rapid onset of action. Neuron. 2007;55(5):712–725. doi:10.1016/j.neuron.2007.07.041
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.