")
Back to Journals » Psychology Research and Behavior Management » Volume 18
Fear-Pleasure Paradox in Recreational Fear: Neural Correlates and Therapeutic Potential in Depression
Received 18 March 2025
Accepted for publication 14 June 2025
Published 27 June 2025 Volume 2025:18 Pages 1509—1518
DOI https://doi.org/10.2147/PRBM.S529004
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Mei-Chun Cheung
Yuting Zhan,1 Xu Ding2
1Department of Psychology, School of Education and Teach, Ningxia University, Yinchuan City, Ningxia Province, 750021, People’s Republic of China; 2School of Nursing, Shandong Academy of Medical Sciences, Shandong First Medical University, Tai’an City, Shandong Province, 271016, People’s Republic of China
Correspondence: Xu Ding, Email [email protected]
Background: Recreational fear, voluntary exposure to frightening stimuli in safe contexts (eg, horror films), elicits both distress and pleasure. Although paradoxical enjoyment of fear has been documented in healthy populations, its neural and psychological mechanisms, and potential therapeutic value for depression, remain unclear.
Methods: In a mixed‐methods design, Study 1 (N = 216) assessed psychological and physiological responses to standardized fear stimuli across a continuum of depressive symptoms, using heart rate variability, salivary cortisol, and validated self‐report measures. Study 2 (N = 84) employed functional MRI to characterize brain activation during and after exposure to the same stimuli in individuals with mild‐to‐moderate depression.
Results: An inverted‐U‐shaped curve linked fear intensity to enjoyment, with depression severity significantly moderating this relationship (β = – 0.42, p < 0.001): higher symptom levels required stronger stimuli for peak pleasure. Depressed participants showed greater ventromedial prefrontal cortex activation and attenuated amygdala responses, suggesting enhanced engagement of regulatory circuits. fMRI analyses also revealed transient normalization of default‐mode and salience‐network connectivity following fear exposure (t(83) = 3.87, p < 0.001, d = 0.84).
Conclusion: Controlled recreational fear may transiently engage emotion‐regulatory networks and modify maladaptive connectivity patterns in depression, offering a novel adjunctive strategy. These findings are preliminary and correlational; future studies should examine causal effects and long‐term clinical impact.
Keywords: recreational fear, depression, emotion regulation, neuroimaging, default mode network, salience network, controlled exposure
Introduction
The fear-pleasure paradox of recreational fear-the voluntary engagement with frightening stimuli for entertainment or pleasure-has long fascinated psychologists studying human emotion and behavior.1,2 While fear typically functions as an adaptive mechanism to promote avoidance of threatening situations, recreational fear experiences such as horror media, extreme sports, and haunted attractions remain popular across cultures and age groups.3 This apparent contradiction has prompted researchers to investigate the mechanisms that allow fear and enjoyment to coexist. Depression represents a particularly compelling context for examining recreational fear experiences due to its characteristic alterations in emotional processing. Depression, characterized by persistent negative affect, anhedonia, and emotion regulation difficulties,4 fundamentally alters how individuals experience and respond to emotional stimuli. Individuals with depression often exhibit altered emotional processing, including blunted positive emotional responses and heightened negative emotional persistence.5 However, despite the potential implications for understanding emotion regulation mechanisms and developing novel therapeutic approaches, the relationship between depression and recreational fear experiences remains largely unexplored.
Recent neuroimaging research has identified specific neural circuit dysfunctions in depression that may be particularly relevant to understanding recreational fear responses. Neuroimaging studies have identified that dysfunctional connectivity between the DMN and SN is often impaired.6,7 The default mode network, associated with self-referential thinking and rumination, shows hyperconnectivity with the salience network in depression, contributing to persistent negative self-focus and difficulty disengaging from distressing thoughts.8 Recent neuroimaging studies continue to underscore aberrant DMN--SN connectivity in depression and even suggest that intense emotional experiences could modulate these networks.9 Furthermore, individuals with depression show deficits in fear extinction processes,10 which involves the ventromedial prefrontal cortex (vmPFC), a region critical for emotion regulation and safety learning.11 These considerations suggest that neural signatures of depression may be temporarily altered during intense emotional experiences that demand attention and presence, such as recreational fear.12 Building on established theories of emotion regulation, we propose a theoretical framework for understanding how recreational fear might influence depressive symptomatology. The potential therapeutic value of recreational fear experiences stems from their unique combination of characteristics: they are voluntary, controlled, time-limited, and occur within safe contexts while eliciting genuine physiological fear responses.13 This combination aligns with principles from Gross’s process model of emotion regulation, which emphasizes the importance of context and cognitive appraisal in emotional responses.14 Additionally, the controlled nature of recreational fear parallels fear extinction paradigms in which safety signals are present alongside threat cues, potentially engaging similar neural mechanisms.15 These features may facilitate engagement with fear stimuli in ways that differ fundamentally from pathological anxiety or traumatic experiences. Furthermore, successful navigation of recreational fear may provide opportunities for mastery experiences and emotion regulation skill development.16 Recent work by Andersen et al17 has demonstrated that enjoyment during recreational fear experiences follows an inverted-U-shaped relationship with fear intensity, suggesting an optimal level of arousal for maximum enjoyment. This pattern aligns with classic psychological theories of optimal arousal18 and flow experiences19 which propose that engagement and enjoyment are maximized when challenges are balanced with perceived abilities to cope. For individuals with depression, who often experience emotional context insensitivity and anhedonia,20 exposure to calibrated fear stimuli may temporarily restore emotional responsiveness and promote engagement with the present moment.
We propose a dual-process framework (rather than “cognitive duality”) wherein recreational fear experiences simultaneously engage threat-processing and safety-signaling neural circuits, creating a unique state of “controlled arousal” that may temporarily disrupt depressive cognitive-affective patterns. This idea aligns with established emotion regulation theories (specifically Gross’s process model of emotion regulation [26]) and with fear-conditioning research on concurrent threat and safety cues, providing a theoretical basis for the framework. Specifically, we hypothesize that recreational fear experiences engage emotion-regulation networks while simultaneously activating reward-processing systems, potentially counteracting the anhedonia and emotional blunting characteristic of depression. We focused particularly on the vmPFC due to its established role in fear extinction and emotion regulation,11 and on DMN-SN connectivity given that DMN-SN hyperconnectivity is strongly linked to rumination in depression.8 To address critical gaps in the literature, this research addresses three primary questions:
- How do individuals with varying levels of depressive symptoms respond psychologically and physiologically to recreational fear experiences? This question addresses the lack of systematic investigation into how depression severity modulates responses to recreational fear.
- What neural mechanisms underlie these responses? Understanding these mechanisms is critical for determining whether any observed effects represent meaningful alterations in brain function that could have therapeutic relevance.
- Could exposure to recreational fear be associated with temporary changes in dysfunctional neural activity patterns associated with depression? This question directly examines the therapeutic potential by investigating whether recreational fear can influence the specific neural circuits implicated in depression.
It is important to acknowledge potential limitations and risks of this approach. It should be noted, however, that recreational fear may not be universally beneficial-intense fear stimuli could exacerbate anxiety or distress in certain vulnerable individuals (those with severe depression or trauma histories). Individuals with comorbid anxiety disorders, post-traumatic stress disorder, or those with severe depression may experience increased distress rather than benefit from fear exposure.22 Additionally, prior exposure to horror media and individual differences in fear tolerance may significantly influence responses.23 To preempt such concerns, we emphasize that our approach is exploratory and should be applied cautiously, with appropriate screening and ethical safeguards.
Methods
Participants
Participants were recruited through university research participant pools, community advertisements, and social media platforms. For Study 1, 216 adults (120 female, mean age = 28.6 years, SD = 7.2, range = 18–45) were enrolled. Participants were categorized based on Patient Health Questionnaire (PHQ-9) scores as having minimal (0–4, n = 72), mild (5–9, n = 63), moderate (10–14, n = 49), or moderately severe to severe (15+, n = 32) depressive symptoms. Exclusion criteria included current psychotic disorders, bipolar disorder, substance use disorders, history of cardiovascular disease, pregnancy, and use of beta-blockers or benzodiazepines. For Study 2, 84 participants (46 female, mean age = 29.4 years, SD = 6.8) with mild-to-moderate depression (PHQ-9 scores 10--14) were included. Participants with severe depression (PHQ-9 ≥ 15) were excluded for both ethical and practical reasons: to ensure participant safety during fear exposure (as severe depression often involves heightened vulnerability to distress) and because severe depression typically necessitates immediate clinical intervention rather than research participation. Additional requirements included no current psychotherapy or antidepressant medication use in order to avoid confounding effects on neural responses. Additional exclusion criteria included standard MRI contraindications (metal implants, claustrophobia, etc). All participants provided written informed consent. The study was approved by the institutional review boards of all participating institutions and adhered to all relevant guidelines and regulations.
Study 1: Psychological and Physiological Assessment
Participants completed baseline psychological measures including the PHQ-9,21 Difficulties in Emotion Regulation Scale (DERS;22), Behavioral Inhibition/Behavioral Activation Scales (BIS/BAS;23), and Sensation Seeking Scale.11 Heart rate monitoring was conducted using Firstbeat Bodyguard 2 devices (Firstbeat Technologies, Finland), and salivary cortisol samples were collected at five time points (baseline, pre-exposure, immediate post-exposure, 20 minutes post-exposure, and 40 minutes post-exposure). Participants were then exposed to a 20-minute guided tour of a professionally designed haunted attraction featuring six standardized fear-inducing scenarios with increasing intensity. Detailed descriptions of the haunted attraction design, including environmental controls, actor behaviors, and safety procedures, are provided in the Table S1, details in the Supplementary Material. After each scenario, participants completed brief assessments of fear intensity and enjoyment using visual analog scales (0–100). Post-exposure, participants completed comprehensive measures of emotional responses, perceived control, and subjective experience. Participants in Study 1 were exposed to standardized horror media clips and a controlled haunted house scenario. Detailed descriptions of the horror film clips (including titles, durations, and intensity ratings from pilot testing) and the design of the haunted attraction scenario are provided in Supplementary Material.
Study 2: Neuroimaging Protocol
Participants underwent fMRI scanning using a 3T Siemens Prisma scanner with a 64-channel head coil. After structural scans (T1-weighted MPRAGE: TR = 2300ms, TE = 2.32ms, flip angle = 8°, 1mm isotropic voxels), participants completed three functional scanning sessions: baseline resting-state (8 minutes), exposure to horror stimuli (12 minutes), and post-exposure resting-state (8 minutes). During the horror stimuli exposure, participants viewed a series of 30-second clips from professionally produced horror media interspersed with 15-second neutral segments. Horror stimuli were selected from mainstream films with PG or PG-13 ratings, focusing on psychological suspense and jump scares while avoiding graphic violence or disturbing imagery. Specific clip details and pilot-tested fear intensity ratings are provided in Supplementary Material. Participants rated fear and enjoyment after each clip using a button-box (1–5 scale). Heart rate was monitored continuously throughout the scanning session using an MRI-compatible pulse oximeter.
Functional images were acquired using a multiband echo-planar imaging sequence (TR = 720ms, TE = 33ms, flip angle = 52°, multiband factor = 8, 2 mm isotropic voxels, 72 slices). Field maps and physiological monitoring data (respiration, cardiac) were acquired for subsequent correction of artifacts. All sessions were conducted under standardized conditions (low ambient lighting, controlled sound levels) to ensure a consistent fear-induction setting for all participants.
Data Analysis
Behavioral and Physiological Data
Self-reported fear and enjoyment ratings were analyzed using hierarchical linear modeling with participants as random effects. We tested linear and quadratic models to examine the relationship between fear intensity and enjoyment, with depression severity as a moderating variable. Physiological data (heart rate, HRV, cortisol) were preprocessed to remove artifacts and normalize distributions before statistical analysis. Time-domain (RMSSD, SDNN) and frequency-domain (LF, HF, LF/HF ratio) HRV measures were calculated. Heart rate variability (HRV) signals were filtered using a 5-point moving average filter, and outlier intervals exceeding 3 standard deviations from the mean were excluded from analysis to remove ectopic beats and artifacts.
Neuroimaging Analysis
Functional MRI data were preprocessed using fMRIPrep 20.2.0 (Esteban et al, 2019) with the following steps: motion correction, slice timing correction, distortion correction using field maps, registration to structural images, normalization to MNI space, spatial smoothing (6mm FWHM Gaussian kernel), and ICA-based denoising. Scans with motion exceeding 3 mm translation or 3 degrees rotation were excluded from analysis. Physiological noise removal was performed using aCompCor. Task-based analyses employed a general linear model (GLM) approach with fear intensity ratings as parametric modulators. Region of interest (ROI) analyses focused on the amygdala, insula, ventromedial prefrontal cortex (vmPFC), dorsolateral prefrontal cortex (dlPFC), and anterior cingulate cortex (ACC). Resting-state functional connectivity analyses compared pre- and post-exposure scans, focusing on connectivity between nodes of the DMN (medial prefrontal cortex, posterior cingulate cortex), SN (anterior insula, dorsal ACC), and CEN (dlPFC, posterior parietal cortex). Network analysis utilized graph theoretical approaches to assess global and local efficiency, modularity, and participation coefficient metrics.
Results
Relationship Between Fear Intensity and Enjoyment
Consistent with previous research, we observed an inverted-U-shaped relationship between fear intensity and enjoyment across all participants (β = −0.31, SE = 0.07, p <0.001). However, this relationship was significantly moderated by depression severity (interaction term: β = −0.42, SE = 0.09, p <0.001). As shown in Figure 1, participants with minimal depression exhibited the strongest quadratic relationship, while those with moderately severe to severe depression showed a flattened response curve, indicating reduced differentiation between fear intensity levels. Interestingly, participants with moderate depression showed a distinctive pattern wherein their peak enjoyment occurred at higher fear intensity levels compared to those with minimal or mild depression (F(3,212) = 8.76, p <0.001, η² = 0.11). This suggests a potential “optimal arousal threshold shift” wherein individuals with moderate depression may require greater stimulation to achieve peak enjoyment, possibly reflecting the emotional blunting often observed in depression.
 |
Figure 1 Relationship between fear intensity and enjoyment across depression severity groups. This figure illustrates the inverted-U-shaped relationship between fear intensity and enjoyment, with notable differences across depression severity groups. The minimal depression group shows the strongest quadratic relationship with peak enjoyment at moderate fear levels. The moderate depression group demonstrates a rightward shift in peak enjoyment (requiring higher fear intensity), while the moderately severe/severe depression group exhibits a flattened response curve, indicating reduced differentiation between fear intensity levels. |
Physiological Responses
Analysis of heart rate variability revealed significant group differences in autonomic nervous system regulation during fear exposure. Table 1 presents the HRV metrics across depression severity groups, demonstrating progressively reduced vagal tone (RMSSD and HF) with increasing depression severity. Cortisol responses showed a significant group × time interaction (F(12,636) = 3.86, p <0.001, η² = 0.07), with the moderate depression group showing the most pronounced cortisol reactivity and recovery pattern. This finding aligns with the psychological data suggesting that moderate depression may be associated with a state of optimal responsiveness to recreational fear experiences.
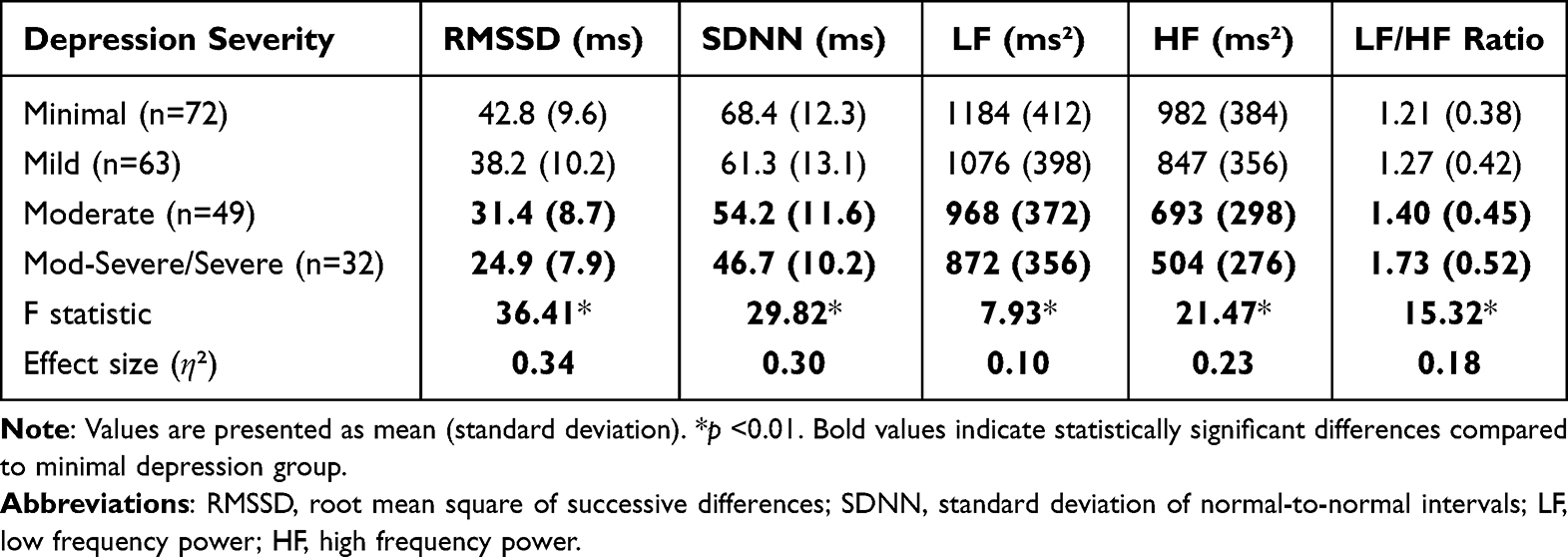 |
Table 1 Heart Rate Variability Measures Across Depression Severity Groups |
Neural Activation Patterns
Task-based fMRI analyses revealed distinct neural activation patterns during recreational fear exposure. Across all participants, fear stimuli elicited increased activation in regions associated with threat processing (amygdala, insula) and emotional regulation (vmPFC, dlPFC). However, when examining activations as a function of subjective fear ratings, we observed significant differences between participants based on depression severity. Particularly noteworthy was the pattern observed in vmPFC activation. As shown in Figure 2, participants with moderate depression showed stronger vmPFC recruitment during peak fear experiences compared to matched non-depressed controls (t(82) = 3.42, p =0.001, d = 0.75). This enhanced recruitment of emotion regulation regions may represent a compensatory mechanism in response to fear stimuli.
 |
Figure 2 Neural activation patterns during recreational fear exposure. This three-part figure shows: (A) A sagittal view of the brain highlighting ventromedial prefrontal cortex (vmPFC) activation during peak fear experiences; (B) A comparison showing significantly higher vmPFC activation in the moderate depression group versus matched controls (t(82) = 3.42, p =0.001, d = 0.75); (C) A positive correlation between vmPFC activation and subjective enjoyment ratings (r = 0.38, p <0.001). The moderate depression group demonstrated stronger vmPFC recruitment during fear stimuli compared to matched controls, potentially indicating compensatory regulatory mechanisms. Significant positive correlation between vmPFC activation and subjective enjoyment ratings suggests that successful regulation of fear may promote positive emotional experiences, potentially counteracting anhedonia. The Figure 2 artwork contains an asterisk indicator (e.g**) means p <0.001. |
Network Connectivity Analysis
Comparison of pre- and post-exposure resting-state scans revealed significant alterations in functional connectivity patterns following recreational fear exposure. Of particular interest were changes in DMN-SN connectivity, which showed temporary alterations in depressed participants following the fear experience. As illustrated in Figure 3, participants with moderate depression exhibited hyperconnectivity between posterior DMN nodes and key salience network regions at baseline, a pattern associated with ruminative processes in depression. Following the fear experience, this hyperconnectivity was temporarily reduced to levels comparable to non-depressed controls (t(83) = 3.87, p <0.001, d = 0.84). This effect persisted through the 8-minute post-exposure scan but began to return toward baseline levels by the end of the session, indicating the temporary nature of these changes. Graph theoretical analyses further supported the observation of network reconfiguration following fear exposure. Global efficiency increased (t(83) = 2.94, p =0.004, d = 0.64), while modularity decreased (t(83) = −3.18, p =0.002, d = 0.69), suggesting a shift toward more integrated neural processing. Moreover, participation coefficient analyses indicated that key SN nodes (particularly the right anterior insula) demonstrated increased between-network connectivity following fear exposure.
 |
Figure 3 Changes in DMN to SN connectivity following recreational fear exposure. This figure presents: (A) Functional connectivity matrices displaying DMN-SN connections before and after fear exposure, with reduced connectivity (lighter colors) in the post-exposure condition; (B) A time course showing temporary reduction of hyperconnectivity in the moderate depression group following fear exposure, with statistical significance (t(83) = 3.87, p <0.001, d = 0.84); (C) A positive correlation between reduction in DMN-SN hyperconnectivity and enjoyment ratings (r = 0.41, p <0.001). **p <0.001. Abbreviations: DMN, default mode network; SN, salience network; PCC, posterior cingulate cortex; mPFC, medial prefrontal cortex; IPL, inferior parietal lobule; MTG, middle temporal gyrus; dACC, dorsal anterior cingulate cortex; AI, anterior insula; SMG, supramarginal gyrus; L, left; R, right. |
Correlation Between Neural and Psychological Measures
Integration of neural and psychological data revealed significant correlations between network connectivity changes and subjective experience measures. Reduction in DMN-SN hyperconnectivity was positively correlated with enjoyment ratings (r = 0.41, p <0.001) and negatively correlated with rumination (r = −0.36, p =0.001). Additionally, vmPFC activation during fear stimuli was positively associated with post-exposure positive affect (r = 0.38, p <0.001). Multiple regression analysis indicated that the combination of vmPFC activation during fear stimuli and post-exposure reduction in DMN-SN connectivity explained 34% of the variance in enjoyment ratings among participants with moderate depression (F(2,81) = 20.85, p <0.001, R² = 0.34).
Discussion
The present study investigated the psychological and neural mechanisms underlying recreational fear experiences in the context of depression, revealing complex relationships that advance our understanding of emotion processing in both normative and clinical populations. Our findings demonstrate that while the relationship between fear intensity and enjoyment follows an inverted-U-shaped curve across participants, this relationship is significantly moderated by depression severity. The observed “optimal arousal threshold shift” in moderately depressed individuals suggests that they may require higher levels of stimulation to achieve peak enjoyment, potentially reflecting the emotional blunting characteristic of depression.24 From a neural perspective, our findings provide evidence for temporary changes in dysfunctional connectivity patterns following recreational fear exposure. Hyperconnectivity between the DMN and SN, which is associated with rumination and negative self-focus in depression,8 was temporarily altered following fear exposure. However, it is important to note that these changes were correlational and temporary in nature, returning toward baseline by the end of the scanning session. This observation aligns with emerging research suggesting that intense emotional experiences can influence maladaptive neural patterns and potentially create windows of opportunity for therapeutic intervention.25–27 The increased recruitment of vmPFC during fear processing among moderately depressed participants represents a particularly intriguing finding. The vmPFC plays a critical role in fear extinction and emotion regulation,11 and its enhanced activation during recreational fear may reflect compensatory regulatory efforts. Furthermore, the correlation between vmPFC activation and post-exposure positive affect suggests that successful regulation of fear in a controlled context may promote positive emotional experiences, potentially counteracting anhedonia. These findings can be integrated within our proposed dual-process framework, wherein recreational fear experiences simultaneously engage threat-processing and safety-signaling neural circuits. This dual activation creates a unique state of controlled arousal that may temporarily influence depressive patterns by:
- Redirecting attention from internal rumination to external stimuli through salience network engagement.
- Activating emotion regulation networks through controlled exposure to fear stimuli.
- Providing opportunities for mastery experiences and positive reinforcement.
- Temporarily altering dysfunctional neural connectivity patterns.
Our results have several theoretical and practical implications. From a theoretical perspective, they extend existing models of emotion processing in depression by highlighting the potential role of controlled arousal in temporarily modifying emotional responses. This aligns with dimensional models of psychopathology that emphasize the role of positive and negative valence systems in affective disorders.25 From a practical standpoint, our findings suggest potential therapeutic applications of recreational fear experiences as adjunctive approaches to conventional depression treatments. However, these applications remain highly speculative and require extensive further research to establish safety and efficacy.
Several important limitations should be considered when interpreting these results. First, our sample included primarily individuals with mild-to-moderate depression, limiting generalizability to those with more severe depression. Additionally, we did not control for individual differences in prior horror media exposure, fear tolerance, or comorbid anxiety symptoms, all of which may significantly influence responses to recreational fear. Second, the laboratory and controlled haunted attraction settings may not fully capture the complexity of naturalistic recreational fear experiences. Third, we did not include a high-arousal but non-fear control condition (such as exciting but non-frightening stimuli), making it unclear whether the observed effects are specific to fear-induced arousal versus general emotional arousal. Fourth, while we observed temporary changes in neural patterns, the correlational nature of our design precludes causal inferences, and the duration of these effects and their potential clinical significance require further investigation through longitudinal studies. Our findings demonstrate temporary reductions in dysfunctional DMN-SN connectivity patterns following recreational fear exposure, but we cannot determine whether these changes causally influence mood or represent epiphenomena of the fear experience.
Future research should examine individual difference factors that may predict responsiveness to recreational fear experiences, explore the durability of the observed neural changes through longitudinal designs, investigate potential therapeutic applications through randomized controlled trials, and examine the effects of different types of recreational fear stimuli and various fear intensities. Most importantly, establishing therapeutic efficacy will require studies that determine optimal exposure parameters and comprehensive safety protocols, particularly for identifying which patients might benefit versus those who might be at risk for adverse effects.
In conclusion, our findings reveal that recreational fear experiences are associated with complex changes indepressive symptomatology and corresponding alterations in both psychological responses and neural connectivity patterns. The dual-process framework proposed in this study offers a novel perspective on the fear-pleasure paradox and its potential implications for understanding emotion regulation in depression. However, any clinical applications remain highly speculative and require extensive further research to establish safety, efficacy, and appropriate implementation protocols. As we continue to explore the neural basis of emotional experiences in clinical populations, recreational fear provides a unique window into the dynamic interplay between negative affect, positive engagement, and neural network function.
Data Sharing Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics Approval and Consent to Participate
The study was reviewed and approved by the Ethics Committee of the Shandong First Medical University. The committee verified that all methods used in this study were carried out in accordance with the 1964 helsinki declaration and its subsequent revisions or similar ethical standards, as well as the ethical requirements of the institutional research committee. Informed consent was obtained from all participants prior to their involvement in this study.
Acknowledgments
The authors would like to thank all participants who took part in this study. We also thank the anonymous reviewers for their valuable comments and suggestions that helped improve this paper.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
There is no funding to report.
Disclosure
The authors declare that they have no competing interests.
References
1. Andersen MM, Schjoedt U, Price H, Rosas FE, Scrivner C, Clasen M. Playing with fear: a field study in recreational horror. Psychol Sci. 2020;31(12):1497–1510. doi:10.1177/0956797620972116
2. Berlyne DE. Conflict, Arousal, and Curiosity. McGraw-Hill; 1960. doi:10.1037/11164-000
3. Bylsma LM, Morris BH, Rottenberg J. A meta-analysis of emotional reactivity in major depressive disorder. Clinic Psychol Rev. 2008;28(4):676–691. doi:10.1016/j.cpr.2007.10.001
4. Carver CS, White TL. Behavioral inhibition, behavioral activation, and affective responses to impending reward and punishment: the BIS/BAS Scales. J Personality Social Psychol. 1994;67(2):319–333. doi:10.1037/0022-3514.67.2.319
5. Clasen M, Kjeldgaard-Christiansen J, Johnson JA. Horror, personality, and threat simulation: a survey on the psychology of scary media. Evol Behav Sci. 2020;14(3):213–230. doi:10.1037/ebs0000152
6. Etkin A, Büchel C, Gross JJ. The neural bases of emotion regulation. Nat Rev Neurosci. 2015;16(11):693–700. doi:10.1038/nrn4044
7. Gratz KL, Roemer L. Multidimensional assessment of emotion regulation and dysregulation: development, factor structure, and initial validation of the difficulties in emotion regulation scale. J Psychopathol Behav Assessment. 2004;26(1):41–54. doi:10.1023/B:JOBA.0000007455.08539.94
8. Williams LM. Precision psychiatry: a neural circuit taxonomy for depression and anxiety. Lancet Psychiatry. 2016;3(5):472–480. doi:10.1016/S2215-0366(15)00579-9
9. Satpute AB, Lindquist KA The default mode network’s role in discrete emotion. Trends Cognit Sci. 2019;23(10):851–864. doi:10.1016/j.tics.2019.07.003
10. Wurst C, Schiele MA, Stonawski S, Weiß C, Nitschke F Impaired fear learning and extinction, but not generalization, in anxious and non-anxious depression. J psychiatric Res. 2021;135:294–301. doi:10.1016/j.jpsychires.2021.01.034
11. Rottenberg J. Mood and emotion in major depression. Curr Directions Psychol Sci. 2005;14(3):167–170. doi:10.1111/j.0963-7214.2005.00354.x
12. Gross JJ, Thompson RA. Emotion regulation: conceptual foundations. In: Gross JJ, editor. Handbook of Emotion Regulation. Guilford Press; 2007:3–24.
13. Hamilton JP, Farmer M, Fogelman P, Gotlib IH. Depressive rumination, the default-mode network, and the dark matter of clinical neuroscience. Biol Psychiatry. 2015;78(4):224–230. doi:10.1016/j.biopsych.2015.02.020
14. Gross JJ The emerging field of emotion regulation: an integrative review. Rev Gen Psychol. 1998;2(3):271–299.
15. Milad MR, Quirk GJ Neurons in medial prefrontal cortex signal memory for fear extinction. Nature. 2002;420(6911):70–74.
16. Hendrikx LJ, van Tol MJ, van der Wee NJA, Schene AH. Potential for neural effects of psychological treatments for depression: a systematic review and meta-analysis. Neurosci Biobehav Rev. 2022;138:104679. doi:10.1016/j.neubiorev.2022.104679
17. Insel T, Cuthbert B, Garvey M, et al. Research Domain Criteria (RDoC): toward a new classification framework for research on mental disorders. Am J Psychiatry. 2010;167(7):748–751. doi:10.1176/appi.ajp.2010.09091379
18. Kaiser RH, Andrews-Hanna JR, Wager TD, Pizzagalli DA. Large-scale network dysfunction in major depressive disorder: a meta-analysis of resting-state functional connectivity. JAMA Psychiatry. 2015;72(6):603–611. doi:10.1001/jamapsychiatry.2015.0071
19. Kerr M, Siegle GJ, Orsini J. Voluntary arousing negative experiences (VANE): why we like to be scared. Emotion. 2019;19(4):682–698. doi:10.1037/emo0000470
20. Lynch T, Martins N. Nothing to fear? An analysis of college students’ fear experiences with video games. J Broadcasting Electronic Media. 2015;59(2):298–317. doi:10.1080/08838151.2015.1029128
21. Kroenke K, Spitzer RL, Williams JB. The PHQ-9: validity of a brief depression severity measure. J Gen Intern Med. 2001;16(9):606–613. doi:10.1046/j.1525-1497.2001.016009606.x
22. Menon V. Large-scale brain networks and psychopathology: a unifying triple network model. Trends Cognitive Sci. 2011;15(10):483–506. doi:10.1016/j.tics.2011.08.003
23. Mobbs D, Yu R, Rowe JB, Eich H, FeldmanHall O, Dalgleish T. Neural activity associated with monitoring the oscillating threat value of a tarantula. Proc Natl Acad Sci. 2010;107(47):20582–20586. doi:10.1073/pnas.1009076107
24. Rottenberg J, Gross JJ, Gotlib IH. Emotion context insensitivity in major depressive disorder. J Abnormal Psychol. 2005;114(4):627–639. doi:10.1037/0021-843X.114.4.627
25. Zuckerman M. Behavioral Expressions and Biosocial Bases of Sensation Seeking. Cambridge University Press; 1994.
26. Csikszentmihalyi M. Flow: The Psychology of Optimal Experience. Harper & Row; 1990.
27. Esteban O, Markiewicz CJ, Blair RW, et al. fMRIPrep: a robust preprocessing pipeline for functional MRI. Nat Methods. 2019;16(1):111–116. doi:10.1038/s41592-018-0235-4
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.