")
Back to Journals » Journal of Inflammation Research » Volume 18
Host Immune Response to Mycobacterium tuberculosis Infection: Implications for Vaccine Development
Authors Liu Q, Que S, Qiu Y, Tang M, Liu S, Yang G, Wang Y, Deng A, Hu X, Lian X, Gao Q
Received 11 January 2025
Accepted for publication 31 May 2025
Published 27 June 2025 Volume 2025:18 Pages 8429—8445
DOI https://doi.org/10.2147/JIR.S517034
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Qianfei Liu,1,2 Songnian Que,1,2 Yixuan Qiu,1,2 Mingxing Tang,1,2 Shaohua Liu,1,2 Guanteng Yang,1,2 Yuxiang Wang,1,2 Ang Deng,1,2 Xiaojiang Hu,3 Xuehui Lian,4,* Qile Gao1,2,*
1Department of Spine Surgery and Orthopaedics, Xiangya Hospital, Central South University, Changsha, Hunan, People’s Republic of China; 2National Clinical Research Center for Geriatric Disorders, Xiangya Hospital, Central South University, Changsha, Hunan, People’s Republic of China; 3Department of Orthopedics, The Second Xiangya Hospital of Central South University, Changsha, Hunan, People’s Republic of China; 4Department of Spine Surgery, The First People’s Hospital of Guiyang City, Guiyang, Guizhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Qile Gao, Department of Spine Surgery and Orthopaedics, Xiangya Hospital, Central South University, Changsha, Hunan, People’s Republic of China, Email [email protected] Xuehui Lian, Department of Spine Surgery, The First People’s Hospital of Guiyang City, Guiyang, Guizhou, People’s Republic of China, Email [email protected]
Abstract: Tuberculosis (TB) is an infectious disease caused by Mycobacterium tuberculosis (Mtb) infection, including pulmonary tuberculosis and extrapulmonary tuberculosis. About a quarter of the people in the world are infected with TB, but only 5– 10% of them will progress to active TB, posing a major challenge to the eradication of TB. The study of the host immune response to Mtb infection is a key aspect of the development of effective vaccines and immunotherapies to eradicate tuberculosis. In this review, we delve into the overview of animal models of TB infection and the host’s innate and adaptive immune responses to Mtb infection. We discuss how Mtb is recognized and phagocytosed by macrophages, how it evades immune responses, the recruitment and mobilization of neutrophils and monocytes, the role of natural killer cells during the infection process, how dendritic cells initiate adaptive immunity, the important roles of CD4+ T cells and their subtypes in TB infection, how CD8+ T cells exert cytotoxic functions, and how B cells produce antibodies and exhibit memory characteristics to eliminate pathogens. Furthermore, we review the tuberculosis vaccines currently entering clinical trials, emphasizing that studying the host’s immune responses following Mtb infection is crucial for the development of more effective vaccines, providing a theoretical foundation and direction for the treatment of tuberculosis.
Keywords: Mycobacterium tuberculosis, immune response, vaccine, infection
Introduction
Tuberculosis primarily affects the lungs which caused by Mycobacterium tuberculosis (Mtb) infection and is transmitted through airborne particles. According to the World Health Organization (WHO), a global total of 8.2 million people were reported as newly diagnosed with TB in 2023, up from 7.5 million in 2022 and 7.1 million in 2019 and far above the levels of 5.8 million in 2020 and 6.4 million in 2021.1 Mtb is transmitted to the lungs through aerosol. Successful infection relies on various factors, such as the proximity and duration of contact with active tuberculosis (ATB) patients, and the immune function of the host.2 Clinically, Mtb infection manifests as latent tuberculosis infection (LTBI), presenting significant challenges for tuberculosis eradication. Approximately one-quarter of the global population have latent Mycobacterium tuberculosis infection, but only 5–10% of these individuals progress to ATB,3 which is characterized by persistent cough, sputum production, weight loss, fatigue, and night sweats. Investigating the host immune response to Mtb infection is a critical aspect of developing effective vaccines and immunotherapies for the eradication of tuberculosis. Innate immune cells in the lungs, primarily macrophages, neutrophils, monocytes, and dendritic cells (DCs), rapidly engulf Mtb and constitute the first line of defense against the pathogen.4 Adaptive immune cells such as CD4+ T cells, CD8+ T cells, and B cells, are involved in killing bacteria and inhibiting bacterial replication.5 Despite the host immune system’s pressure, Mtb has evolved various strategies to evade and disrupt immune responses, allowing it to persist within the host. The immune response to Mtb infection involves the participation of multiple innate and adaptive immune cells.6 A thorough investigation of the complex interactions between Mtb and the host immune system will aid in identifying new therapeutic targets for tuberculosis and is also of great significance for its prevention.
Based on the animal model of Mtb infection, this review discusses the host innate and adaptive immune responses after Mtb infection, as well as the progress of existing tuberculosis vaccines, emphasizing that the study of host immune responses after Mtb infection is crucial for the development of more effective vaccines.
Application of Animal Models in Mycobacterium tuberculosis Infection Research
The study of immune responses to Mycobacterium tuberculosis has greatly benefited from the development of various animal models, including mice, rabbits, fish, and non-human primates (NHPs), each of which has distinct advantages and limitations.7 Mice have been widely used to model the immune response of hosts to infections with clinical isolates of Mtb, including susceptible and non-susceptible mice, which exhibit differences in their immune responses following infection with Mtb.8 Due to the similarities in brain development processes between humans and rabbits, such as neuronal development, myelination, and microglial function, rabbit models have been used to study meningeal and spinal tuberculosis.9,10 However, rabbits exhibit resistance to Mtb, necessitating the use of large bacterial inocula or more virulent strains in model establishment.11 The establishment of zebrafish models has provided new insights into the formation of Mtb-induced granulomas. The pathological process can be visually monitored, and subcellular imaging of infected phagocytes within live hosts is possible.12 NHPs, such as rhesus or cynomolgus monkeys, can be infected via aerosol or bronchoscopy to the lungs, exhibiting many characteristic granulomas observed in humans and presenting clinical symptoms similar to those in humans. However, NHP models are often constrained by ethical considerations, high costs, and difficulties in genetic manipulation.11 In conclusion, animal models of tuberculosis are crucial tools for studying the disease, as they can replicate the natural course of human tuberculosis and facilitate a better understanding of the infection process, pathogenesis, and immune responses to Mtb. These models enable the testing of new tuberculosis vaccines, evaluating their immunogenicity and protective efficacy, and providing a basis for the development of more effective vaccines.
However, it is important to note that animal models also have certain limitations. Although these models can screen for potentially effective vaccine components, they often fail to reliably reflect the true efficacy in humans when assessing the actual performance of the vaccine. There are significant differences in disease resistance/susceptibility among these models, and thus no single model can capture all aspects of the disease pathogenesis.13 Therefore, during the vaccine development process, it is essential to combine various research methods to enhance the likelihood of clinical translation.
Innate Immunity in Mycobacterium tuberculosis Infection
The innate immune system serves as the body’s first line of defense against Mtb, playing a crucial role in the early response to the pathogen. However, innate immune cells can also serve as niches for bacterial replication, with Mtb employing various strategies to undermine the innate immune response and establish chronic infection. Current research on host innate immunity focuses on several aspects: how Mtb is recognized and engulfed by macrophages, the mechanisms by which the bacteria evade immune responses, the recruitment and mobilization of neutrophils and monocytes, the role of natural killer (NK) cells during infection, and how DCs initiate adaptive immunity (Figure 1 and Table 1).
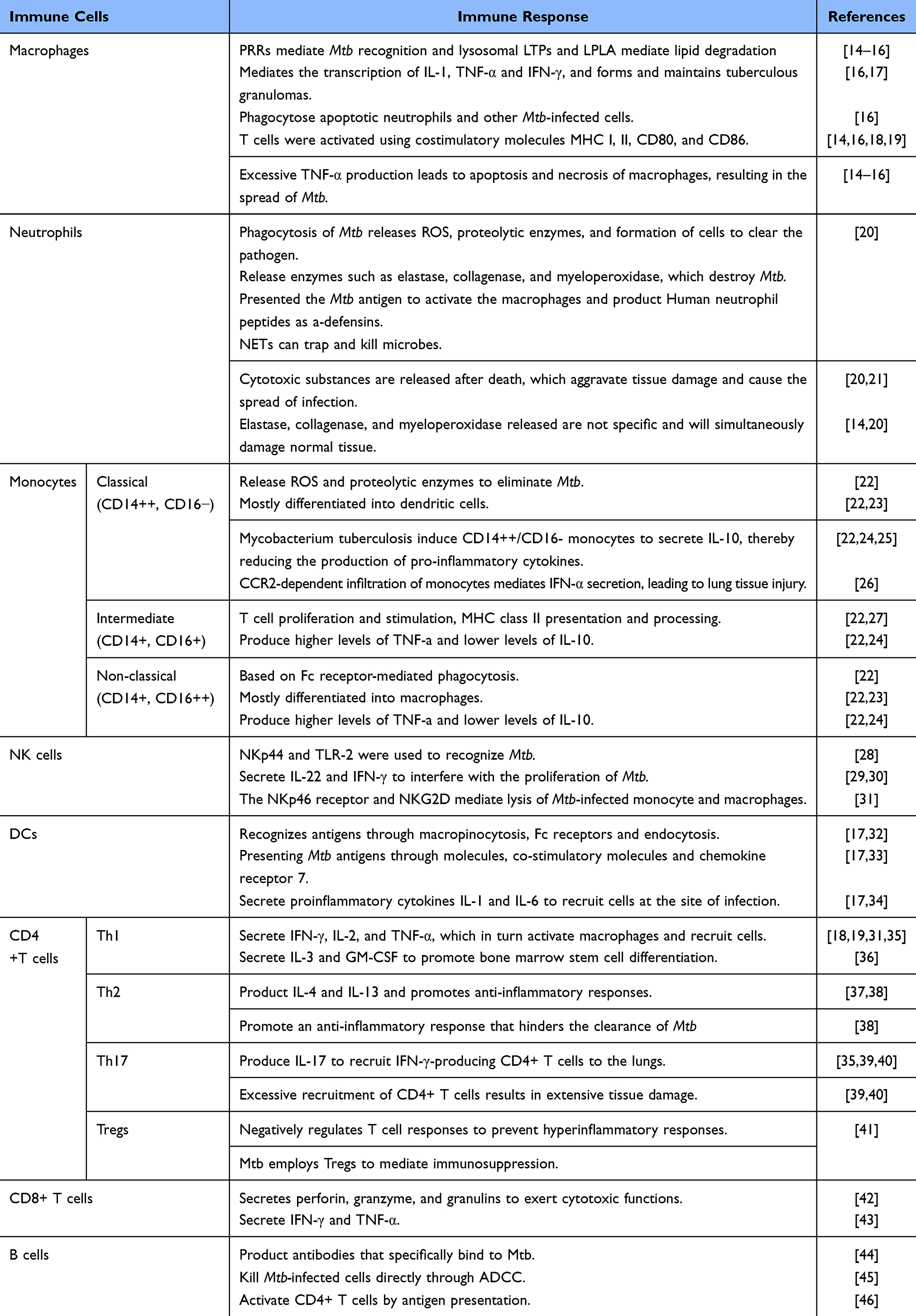 |
Table 1 Strategies of Host Immune Response After Mycobacterium Tuberculosis Infection |
 |
Figure 1 Host immune response after Mycobacterium tuberculosis infection. A) Pattern Recognition Receptors Recognize Mycobacterium tuberculosis; B) Mycobacterium tuberculosis evades the host immune response through various pathways; C) After infection with Mycobacterium tuberculosis, the host initiates innate immunity. Macrophages engulf Mycobacterium tuberculosis, release cytokines, and recruit neutrophils and monocytes. NK cells directly kill infected cells, release cytokines to enhance the immune response; D) Dendritic cells initiate adaptive immunity and reverse-regulate innate immunity. B cells release antibodies to eliminate Mycobacterium tuberculosis. Created with BioRender.com. https://BioRender.com/a29y964. |
Pattern Recognition Receptors Recognize Mycobacterium tuberculosis
Pathogen associated molecular patterns (PAMPs) on Mtb include mannose-capped lipoarabinomannan (ManLAM), phosphatidylinositol mannoside (PIMs), phenolic glycolipids (PGLs), peptidoglycan, Trehalose diester mycolate (TDM), and Phthiocerol dimycocerosates (PDIM), which can be recognized by various cytoplasmic pattern recognition receptors (PRRs), such as C-type lectin receptors (CLRs) (eg, mannose receptor, DC-SIGN, Dectin-1, Dectin-2, Mincle), NOD-like receptors (eg, NOD2), complement receptors (eg, complement receptor 3), scavenger receptors (eg, MARCO, SR-A1, CD36, SR-B1), Fc receptors (eg, FcgR), and Toll-like receptors (TLRs) (TLR-2, TLR-4, TLR-9), thereby inducing the production of downstream cytokines and autophagy.47 For instance, Mtb expresses various TLR ligands, and Toll-like receptors TLR-2, TLR-4, and TLR-9 are involved in host recognition of Mtb.48 Studies have shown that mice lacking the TLR adaptor protein myeloid differentiation factor 88 (MyD88) rapidly succumb to Mtb infection due to insufficient NOS2 expression in MyD88−/− mice, impaired activation of IL-1β or IL-1 receptor (IL-1R) pathways, diminished macrophage responsiveness to interferon-γ signaling,49 and reduced IL-12 and TNF-α responses in macrophages and DCs.50 MyD88 bridges TLR and IL-1 receptor family ligands to IL-1 receptor-associated kinases (IRAKs) in innate immunity, activating multiple downstream pathways including NF-κB, MAPK, and AP1. Investigating the interactions between PAMPs and PRRs and their downstream effects, as well as monitoring specific PAMP levels or PRR activation, may aid in the early detection and diagnosis of tuberculosis. Targeting PRRs or modulating PAMP recognition pathways could potentially lead to the development of more effective vaccines and immunoadjuvants.
Although host recognition of Mtb activates innate immunity, the pathogen has evolved strategies to evade PRR-mediated innate immune responses. Mtb can alter its cell wall components to inhibit host immune recognition and avoid detection by the host immune system. For instance, lipids such as ManLAM in the cell wall can bind to TLR2, activating mast cells to release exosomes and induce macrophage M2 polarization.51 Mtb can also survive within the host by evading autophagy, an important host defense mechanism for eliminating intracellular pathogens. Mtb secretes ESX-1 associated proteins (EspL) to inhibit phagosome maturation and autophagy, thus promoting its survival.52 Additionally, Mtb phosphoribosyl transferase (MtbPRT) inhibits autophagy independently by inducing high histone methylation (H3K9me2/3) on Atg5 and Atg7 promoters through activation of p38-MAPK and EHMT2 methyltransferase-dependent signaling pathways.53 Feng et al found through a gene knockout mouse model that the absence of autophagy genes leads to acute susceptibility to high-dose Mtb infection.54 Research has shown that conditional knockdown of the core autophagy component Atg5 in myeloid cells renders mice extremely susceptible to Mtb, while depletion of other autophagy factors does not affect the infection.55 Mtb evades immune responses by blocking phagosome-lysosome fusion. For instance, Mtb can secrete PknG, which inhibits lysosome maturation by decreasing GlpK and ALD expression while increasing Ag85A and Ag85C expression, thereby enhancing bacterial infectivity, metabolism, growth rate, virulence, and drug resistance.56 Mtb can also secrete proteins such as SapM, which interfere with phagosome maturation and acidification, thereby protecting the bacteria from host enzymatic destruction and enabling Mtb to survive and replicate within macrophages.57 Mtb can also evade immune responses by inducing host cell apoptosis. For example, the lipoprotein ESAT-6 and another secreted protein, tuberculosis necrotizing toxin (TNT), induce necrotic apoptosis, which facilitates the proliferation and dissemination of the bacteria.58 Mtb can evade detection by the immune system, which may make the diagnosis of latent or subclinical infections more difficult. Diagnostic tests for Mtb infection, such as the tuberculin skin test (TST) and interferon-γ release assay (IGRA), depend on the host’s immune response.59 However, if Mtb successfully suppresses this response, these tests may yield false-negative results, leading to missed or delayed diagnoses. Mtb’s immune evasion contributes to its persistent survival within the host, making complete eradication of the bacteria difficult even with prolonged treatment. This persistent survival is associated with an increased risk of drug-resistant tuberculosis, as prolonged antibiotic exposure can lead to the development of resistant mutations in Mtb,60 including intermittent treatment and patient non-compliance with therapy.61 The ability of Mtb to evade immune recognition poses a significant challenge to vaccine development. Traditional vaccines may fail to elicit a sufficiently strong immune response to provide protection, especially in populations with a high burden of latent tuberculosis. Studying Mtb’s immune evasion strategies is crucial for developing new vaccines that can elicit robust and durable immunity.
Macrophages Rely on Phagosomes to Defend Against Mycobacterium tuberculosis
Macrophages are the first immune cells to encounter Mtb during infection and serve as the primary replication niche for this bacterium. Macrophages recognize and engulf Mtb, utilizing lysosomes and phagosomes to eliminate the bacteria.14 Intracellular signaling pathways triggered by PRR ligands induce actin polymerization to form early phagosomes. Research by Kolonko et al indicates that this process requires WASH-driven polymerization; disruption of WASH results in delayed phagosome maturation.62 Within macrophages, phagosomes interact continuously with trans-Golgi transport vesicles, endosomes, and lysosomes, eventually fusing with lysosomes. This process, known as phagosome maturation,63 is marked by acidification of the phagosome lumen mediated by the proton pump vacuolar ATPase (H+-vATPase).64 Ultimately, mature lysosomes mediate the degradation of lipid membranes through lysosomal lipid transfer proteins (LTPs) and lysosomal phospholipase A2 (LPLA2), leading to the clearance of Mtb.15,16 Beyond their phagocytic function, macrophages also play a crucial role in regulating inflammatory responses during Mtb infection. TLRs and CLRs recognize the bacteria and signal through NF-κB, mediating the transcription of cytokines IL-1 and TNF-α. TNF-α secreted by macrophages mediates T cell recruitment and granuloma maintenance following infection.65 However, TNF-α exhibits a dual role in early tuberculosis infection: Clay et al induced TNF-α signaling deficiency in zebrafish, resulting in accelerated Mtb growth and significantly increased granuloma size.66 Research by Roca et al indicates that excessive TNF-α triggers programmed cell death in infected macrophages by producing mitochondrial reactive oxygen species (ROS) and cyclosporin D.67 IL-1 consists of IL-1α and IL-1β, both of which are predominantly secreted by mononuclear macrophages during tuberculosis immunity. A study involving IL-1α and IL-1β gene knockout mice revealed that single-gene knockout mice could still control acute Mtb infection, whereas double-gene knockout mice could not, indicating that both cytokines play a role in combating Mtb infection.68 On the other hand, there are studies indicating that after infection with Mtb, the secretion of type I interferon (IFN-I) by macrophages is significantly increased, which exacerbates macrophage death.69 Moreover, blocking IFN-I signaling enhances the efficacy of the frontline tuberculosis drug rifampicin in mice infected with Mtb.70 At the same time, research by Saqib et al suggests that IFN-I inhibits the uptake of Mtb by neutrophils and drives the recruitment of CD101-negative neutrophils to the lungs, which promotes the extracellular persistence of Mtb, exacerbates epithelial damage, and impairs the production of surfactant.71 During Mtb infection, the increased signaling of IFN-I can prevent T cell proliferation, directly leading to uncontrolled Mtb infection in neutrophils and alveolar macrophages.72 Bacillus Calmette-Guerin (BCG) is currently used clinically for tuberculosis prevention. Recent studies show that hematopoietic stem cells cultured with BCG produce epigenetically modified macrophages, which exhibit significantly enhanced protection against severe Mtb infection compared to non-cultured macrophages.73 This phenomenon, known as trained immunity, differs from adaptive immunity as it is mediated by innate immune cells such as monocytes, macrophages, and NK cells. It has shown beneficial effects against cancer, viral infections, and autoimmune diseases.74 Although there are no current reports on whether trained immunity can fully control Mtb infection in humans, leveraging the modifiability of macrophages to enhance their immune response against Mtb remains a potential avenue for future tuberculosis prevention and treatment.
Recruitment of Neutrophils and Monocytes During Mycobacterium tuberculosis Infection
Following Mtb infection, the recruitment and mobilization of neutrophils constitute a crucial component of the immune response. Neutrophils exhibit a dual role. On one hand, host cells (eg, macrophages and DCs) recognize and engulf pathogens, subsequently releasing various cytokines, such as TNF-α, IL-1β, IL-6, and IL-8. These cytokines not only promote the inflammatory response at the local infection site but also enhance the recruitment and extravasation of neutrophils through the activation of endothelial cells.75 Upon sensing chemokines secreted at the site of inflammation, neutrophils migrate to the infection site by upregulating the expression of chemokine receptors, such as CXCR1 and CXCR2. At the local microenvironment, they perform functions including pathogen phagocytosis and the release of neutrophil extracellular traps (NETs), thereby limiting the spread of Mtb.20,21,76 However, some studies have shown that NETs can promote the extracellular growth of Mtb in mice infected with tuberculosis, leading to pulmonary lesions and inflammation.77 Additionally, cit-H3, apart from being a marker of NETs, is also a potential marker of lung tissue damage induced by severe pulmonary tuberculosis.78–80 On the other hand, excessive activation and persistent presence of neutrophils may lead to tissue damage. This is because the reactive oxygen species and proteases released by neutrophils not only exert lethal effects on pathogens but also damage host tissues. In chronic tuberculosis infections, the sustained recruitment and inflammatory response of neutrophils lead to the formation of localized necrosis and fibrosis, causing structural damage to the lungs and providing a niche environment conducive to Mtb survival.81 A study found that, when comparing C57BL/6 mice resistant to Mtb infection with DBA/2 mice susceptible to it, neutrophils in DBA/2 mice were rapidly recruited to the bronchoalveolar regions and were more numerous. The prolonged clearance of neutrophils in DBA/2 mice at the early stage of infection extended their lifespan, suggesting that the rapid recruitment of neutrophils in genetically susceptible mice is pathogenic.82 However, another study indicated that during the first four days post intravenous injection of Mtb, neutrophil depletion led to enhanced bacterial growth at extrapulmonary sites. This suggests that the protective immunity conferred by neutrophils against Mtb may depend on the route of infection and the dynamics of the infection.83 Due to the dual role of neutrophils in Mtb infection, regulating the recruitment and activation of neutrophils may represent a novel strategy for tuberculosis treatment, particularly in terms of preventing tissue damage and inflammation-related complications.
In addition to neutrophils, monocytes are also recruited to the site of Mtb infection. Based on the expression of surface markers CD14 and CD16, monocytes are primarily categorized into three subsets: classical monocytes (CD14++, CD16−), intermediate monocytes (CD14+, CD16+), and non-classical monocytes (CD14+, CD16++), each with distinct functions.22 Classical monocytes primarily produce reactive oxygen species (ROS). Upon tissue infection or injury, they migrate to the affected tissue and secrete unique chemokines to activate other immune cells. Intermediate monocytes play a crucial role in antigen presentation as well as the secretion of cytokines and pro-inflammatory interleukins. A study indicates that after human infection with Mtb, CD16+ monocytes in peripheral blood are amplified during tuberculosis infection, and the disturbance of this subset determines the severity of the disease.24 Meanwhile, anti-tuberculosis treatment in tuberculosis patients can reverse the amplification of CD16+ monocytes.25 During severe infection, CD16+ monocytes in the peripheral blood of tuberculosis patients upregulate the expression of CC chemokine receptor type 2 (CCR2), aimed at enhancing their migratory capacity to the site of infection.84 Following Mtb infection, monocytes differentiate into macrophages and DCs. Monocytes have been identified as the primary innate immune cell population producing iNOS in Mtb-infected mice.23 Additionally, monocytes deliver Mtb to the pulmonary lymph nodes, where they coordinate with DCs to initiate CD4+ T cell responses against Mtb infection.27 However, monocyte recruitment following Mtb infection may also be detrimental to the host by creating an environment conducive to bacterial growth and replication. Antonelli et al demonstrated that treating Mtb-infected mice with polyinosinic-polycytidylic acid (polyIC) increased the severity and mortality of the infection through a CCR2-mediated mechanism.26
NK Cells in Mycobacterium tuberculosis Infection
NK cells are innate lymphocytes characterized by various surface markers, including CD56, CD16, CD94, NKG2D, and CD57. These cells do not rely on major histocompatibility complex (MHC) molecules to recognize target cells; instead, they identify potential infected cells by detecting “stress molecules” or “missing MHC molecules” on the surface of target cells.85 Various components of the Mtb cell wall can directly bind to NKp44 on NK cells.86 NK cells can also recognize stress molecules upregulated on the surface of Mtb-infected cells, leading to the direct killing of infected macrophages.28 Additionally, NK cells can restrict intracellular bacterial replication through the secretion of IL-2229 and IFN-γ.30 IL-21-activated NK cells further enhance the immune response by increasing the production of IL-1β, IL-18, and MIP-1β. NK cells also lyse Mtb-infected macrophages and alveolar macrophages while upregulating CD8+ T cell responses.31 Interestingly, IL-21-dependent NK cell populations that emerge following BCG vaccination have been shown to expand after Mtb challenge,87 suggesting that NK cells may exhibit some hallmark features of memory cells. Research by Choreño-Parra et al indicates that Mtb antigens stimulate CXCR6+ NK cells in both mice and humans, although their activation mechanisms have not been further evaluated.88 As mentioned previously, innate immune cells such as macrophages, neutrophils, and monocytes are crucial for the early response against Mtb, but they may also serve as niches for bacterial replication to some extent. However, NK cells do not serve as niches for Mtb and do not spread the pathogen. Due to the ability of Mtb to evade immune cell surveillance by suppressing the expression of MHC molecules, NK cells can recognize missing or aberrantly expressed MHC molecules. This characteristic makes them a potential vaccine target. Activating NK cells may help eliminate infected cells that lack MHC molecules, thus effectively controlling tuberculosis infection. Studies have shown that NK cells play an important role in defending against Mycobacterium tuberculosis infection.89 Therefore, a deeper understanding of the recognition mechanisms and functions of NK cells will provide new ideas and strategies for the development of future vaccines.
Dendritic Cells Initiate Adaptive Immunity Against Mycobacterium tuberculosis
During Mtb infection, a critical function of innate immunity is to initiate the adaptive immune response. DCs are specialized antigen-presenting cells that initiate the adaptive immune response by presenting Mtb antigens through (MHC) molecules, co-stimulatory molecules (CD80, CD86, and CD40), and chemokine receptor 7 (CCR7).17 DCs can be categorized into monocyte-derived DCs (moDCs), conventional DCs (cDCs), and plasmacytoid DCs (pDCs) based on surface marker expression.90 However, the functions of these subsets remain controversial and warrant further detailed investigation. Infected DCs migrate to the pulmonary draining lymph nodes, where they secrete soluble, unprocessed Mtb antigens and present them to T cells through MHC molecules. MHC molecules are divided into MHC class I and MHC class II, which present to CD8+ and CD4+ T cells, respectively.91 DCs present antigens to CD4+ T cells with bactericidal capabilities via MHC class II molecules, secreting IFN-γ to activate macrophages.33 Mtb-infected DCs can promote a protective Th17 response against highly virulent Mtb infection by secreting IL-23, IL-6, and IL-1β.34 Cytokines secreted by DCs help regulate the immune response, with surface co-stimulatory molecules binding to corresponding receptors on T cells, thereby activating T cell responses and initiating adaptive immunity. DCs are key participants in initiating the adaptive immune response against Mtb and determining the outcome of infection. Further research on DC cell populations may reveal that interventions or therapies aimed at improving DC function could benefit from enhanced interactions between DCs and antigen-specific T cells. This includes host-directed therapies, development of improved vaccines, and control strategies, which have significant implications for the treatment and prevention of tuberculosis.32
Adaptive Immunity in Mycobacterium tuberculosis Infection
Adaptive immunity refers to the part of the immune system that generates long-lasting, specific responses against particular pathogens. Specific T cells, through cytokine secretion and direct antimicrobial action, are central to the adaptive immune response against Mtb infection. T cells are classified based on surface markers and functions into several types: helper T cells (Th cells), which carry the CD4 molecule on their surface, primarily coordinate immune responses through cytokine secretion. These can be further subdivided into Th1, Th2, Th17, and regulatory T cells (Tregs), with each subset having unique functions based on their cytokine profiles.92 Cytotoxic T cells (CTLs), which carry the CD8 molecule on their surface, directly kill virus-infected or tumor cells.93 CD4+ T cells, CD8+ T cells, and B cells form the basis of humoral and cell-mediated immunity, providing specific responses to various Mtb antigens, which are typically presented by MHC class I and II molecules. However, adaptive immune responses can also have detrimental effects on the host by promoting excessive inflammation or chronic antigen exposure. The following sections will delve into the crucial roles of CD4+ T cells and their subsets in tuberculosis infection, the mechanisms by which CD8+ T cells exert cytotoxic functions, and how B cells produce antibodies and utilize memory characteristics to clear pathogens.
The Role of CD4+ Cells in Mycobacterium tuberculosis Infection
CD4+ T cells play a crucial protective role by secreting a range of cytokines (eg, IFN-γ and TNF-α). These cytokines attract other immune cells to the infection site and promote the differentiation of several CD4+ T cell subsets into effector cells to eliminate Mtb infection.94 Th1 cells activate macrophages by expressing membrane molecules such as CD40L and secreting cytokines like IFN-γ. This activation leads to increased expression of immune molecules such as CD80, CD86, and MHCII on macrophages, and the secretion of cytokines such as IL-12, which in turn enhances Th1 responses.18,19 Additionally, in response to Mtb attack, IL-3 and GM-CSF produced by Th1 cells stimulate the differentiation of hematopoietic stem cells in the bone marrow into monocytes. Meanwhile, cytokines such as TNF-α, LTα, and MCP-1 induce endothelial cells to express more adhesion molecules.36 This process not only facilitates the adhesion of monocytes and lymphocytes to endothelial cells but also assists these cells in traversing the vascular wall and migrating to the infection site. Thus, Th1 cells effectively activate and enhance macrophage function, and during the response to Mtb, they promote the recruitment and aggregation of macrophages and other immune cells, forming a robust immune defense network.35 Th2 cells play a central role in humoral immunity by producing cytokines such as IL-4 and IL-13, which promote the development of anti-inflammatory responses.95 Mtb can cause a shift in the host immune response from a Th1-dominated reaction to a Th2-dominated response.37 A Th2-dominated response is insufficient to effectively control the growth of Mtb over the long term. Although the involvement of Th2 cells may partially alleviate inflammatory responses, it could also be a key factor contributing to the chronic persistence and difficulty in eradicating tuberculosis.38 Th17 cells not only induce neutrophilic inflammation, which can potentially damage host tissues, but also play a significant role by producing IL-17.35 IL-17 induces chemokines in the lungs during infection, which help recruit IFN-γ-producing CD4+ T cells to the lungs, leading to extensive tissue necrosis and thereby enhancing the immune response.39 The depletion of Th17 cells is considered a potential mechanism for the transition of latent tuberculosis infection LTBI to ATB.40 These findings highlight the dual role of Th17 cells in Mtb infection: on one hand, they may cause tissue damage, while on the other hand, they play a critical role in preventing pathogen spread and disease progression. Regulatory T cells (Tregs), a subset of CD4+ T cells, play a crucial role in negatively regulating T cell responses to prevent excessive inflammatory reactions. In the context of Mtb infection, once Mtb within phagocytes migrates from the lesion to the lymph nodes, it activates and promotes the proliferation of Mtb-specific Tregs. Recent research by Wang et al41 using single-cell RNA sequencing, has revealed an increase in Tregs in the lung lesions of tuberculosis patients. This proliferation significantly impacts the immune system of patients with ATB or LTBI, as Mtb can induce immunosuppression by increasing Treg cell numbers, thereby interfering with CD4+ T cell function and control of the disease. Studying the immune response of CD4+ T cells following Mtb infection and the balance among different cell subsets is crucial for understanding the immunopathogenesis of the disease, improving diagnostic methods, developing vaccines and therapies, and monitoring disease progression.
The Role of CD8+ T Cells in Mycobacterium tuberculosis Infection
During the immune response to Mtb infection, CD8+ T cells exert their cytotoxic function by secreting perforin, granzymes, and granulin, thereby directly killing Mtb-infected host cells.42 Key cytokines produced by CD8+ T cells, such as interferon-γ (IFN-γ) and TNF, play crucial roles in regulating immune responses and inflammation, extending beyond their direct cytotoxic capabilities.43 CD8+ T cells and their inherent cytotoxic mechanisms form pores by releasing perforin and other molecules, delivering cytotoxic molecules to target cells to directly kill or eliminate infected macrophages.96 Recent advancements indicate that specific fragments of granzyme exhibit antimicrobial activity and can inhibit Mtb replication within host cells in vitro, further highlighting the multifaceted and complex roles of CD8+ T cells in the host’s defense against Mtb infection.97 Recent studies using a mouse tuberculosis model98 found that during the interaction between Mtb antigen TB10.4 and CD8+ T cells, TB10.4-specific CD8+ T cells failed to effectively recognize Mtb-infected macrophages. This suggests that the TB10.4 antigen may act as a decoy, masking the effective recognition and response to Mtb-infected macrophages. In contrast, specific CD4+ T cells targeting major epitopes such as Ag85b or ESAT6 have demonstrated the ability to recognize and effectively inhibit the growth of Mtb-infected macrophages, highlighting different mechanisms of immune defense by CD4+ T cells compared to CD8+ T cells in Mtb infection. This difference in mechanisms may reflect specific strategies by Mtb to evade and disrupt host immune responses, thereby limiting the efficacy of CD8+ T cells in host defense mechanisms. This finding not only deepens our understanding of the immune response mechanisms to Mtb infection but also provides valuable insights for future vaccine design and therapeutic strategy development.
B Cells in Mycobacterium tuberculosis Infection
While T cells are central to anti-tuberculosis immune responses, recent studies have highlighted the equally significant role of B cells in Mtb infections. B cells are a crucial component of the adaptive immune system, primarily responsible for antibody production and regulation to eliminate pathogens.99 Although cellular immunity is the primary immune response in tuberculosis, studies have shown a significant increase in the production of specific IgG and IgA antibodies following Mtb infection, indicating the critical role of B cells in combating the infection.100 After tuberculosis infection, B cells can differentiate into memory B cells, which rapidly produce antibodies upon re-exposure to the pathogen, thus enhancing the immune response. B cells are not only producers of antibodies but also secrete various cytokines, such as IL-10 and IL-6, which play crucial roles in regulating immune and inflammatory responses. B cells enhance the body’s antibacterial capacity by producing antibodies that bind to Mtb and facilitate macrophage phagocytosis of the pathogen.44 Additionally, B cells can directly kill infected cells through antibody-dependent cell-mediated cytotoxicity (ADCC).45 The interaction between B cells and T cells is critical in tuberculosis infection. B cells can activate CD4+ T cells through antigen presentation and provide auxiliary signals during the immune response, while cytokines secreted by T cells can also influence B cell activation and antibody production. In tuberculosis infections, B cells may also suppress excessive inflammatory responses by secreting immune-regulatory factors (eg, IL-10), thereby preventing damage to the host’s own tissues.46 The production of antibodies, cytokines, and memory characteristics by B cells is crucial for the response to Mtb. Studying how B cells recognize and respond to this pathogen could lay the experimental foundation for developing vaccines that elicit strong humoral immune responses.
Vaccine Development and Challenges
The only vaccine currently approved for the prevention of tuberculosis is BCG, an attenuated strain of Mycobacterium bovis that provides protection against disseminated tuberculosis and tuberculous meningitis.101 The protective benefit of BCG is based on the priming of antigen-specific naive B and T cells to generate memory B and T cells. When naive lymphocytes receive Mtb cognate antigens, they become activated and undergo clonal expansion, in which the transcriptional profiles of B and T cells are epigenetically regulated and differentiate into effector and memory cells,102 Activated B cells undergo immunoglobulin class switching and somatic hypermutation to produce high-affinity antibodies with multiple effector functions. Most activated T cells express effector molecules such as perforin to exert immune effects,103 and a small proportion of CD8+T cells can retain the epigenetic background for decades, rapidly proliferate and exert immune effects upon pathogen re-invasion104 Training immunity is also an important way for BCG to exert its benefits. Studies have found that BCG mediates the reprogramming and differentiation of bone marrow stem cells, and enhances the ability of macrophages to resist Mtb infection by relying on IFN-γ,105 This effect persists long after BCG is eliminated from the body.73 However, BCG live vaccine can prevent the maturation of phagosomes by blocking expression of the lysosomal glycoprotein LAMP-1 in the phagosome exon.106 However, its efficacy remains a subject of controversy. As of now, approximately 20 tuberculosis vaccine candidates are in clinical evaluation, with 15 of them undergoing clinical trials. These vaccines can be classified into attenuated live vaccines, inactivated vaccines, subunit vaccines, and recombinant live vaccines107 (Table 2).
 |
Table 2 Current Research Progress of Vaccines Entering Clinical Trials |
One example of an attenuated live vaccine is MTBVAC. Preclinical studies have highlighted that MTBVAC induces immunity against ESAT6 and CFP10, showing improved efficacy compared to BCG.108 Phase II clinical trials conducted in South Africa in adults and newborns have also validated its immunogenicity, showing that MTBVAC vaccination induces sustained expression of antigen-specific Th1 cytokines in infants.109 A Phase III clinical trial is underway to evaluate the safety, immunogenicity, and efficacy of MTBVAC in infants born to HIV-positive or HIV-negative mothers.110
Three inactivated vaccines currently known to be in clinical trials include RUTI, DAR-901, and Vaccae. Prabowo et al found that RUTI vaccination enhanced the inhibition of ex vivo Mycobacterium growth and induced a shift in the phenotype of murine monocytes, with a significant increase in non-classical monocytes.111 Phase I clinical trials of RUTI assessed its safety and immunogenicity in healthy volunteers, showing that adverse events were positively correlated with dose.112 A randomized, placebo-controlled Phase II clinical trial conducted in patients with latent tuberculosis evaluated the safety, tolerability, and immunogenicity of RUTI.113 Phase I clinical trials of DAR-901 indicated that it has good tolerability and induces a robust CD4+ T cell immune response compared to BCG.114 When used as an adjunctive therapy, Vaccae has been shown to increase CD4+ T cell counts.115 A Phase III clinical trial evaluated the safety and efficacy of Vaccae in preventing tuberculosis, but the results have not yet been published.
Five subunit vaccines include M72/AS01E, H56:IC31, ID93+GLA-SE, GamTBvac, and AEC/BC02. M72/AS01E has undergone a series of Phase II clinical trials in multiple regions, including India, South Africa, and Taiwan. Overall results indicate that M72/AS01E demonstrates good safety and immunogenicity across different populations.116 Currently, the Bill & Melinda Gates Foundation and Wellcome Trust have announced a $550 million funding commitment to advance Phase III clinical trials of M72/AS01E. If successful, M72/AS01E could become the first new tuberculosis vaccine to be licensed in over a century.117 Phase IIa clinical trial results for H56:IC31 show that it can induce persistent antigen-specific CD4+ T cell responses in both Mtb-infected and uninfected adults, with good safety and tolerability.118 Phase IIa clinical trial results for ID93+ GLA-SE demonstrate that this vaccine induces specific multifunctional CD4+ T cell responses and sustained antibody responses (including IgG1 and IgG3 subclasses). The adverse reactions observed were limited to mild indurations and erythema.119 Phase I and II clinical trial results for GamTBVac conducted in Russia indicate that it has good safety and immunogenicity,120 Phase III clinical trial is currently recruiting. Phase I clinical trials for AEC/BC02 have been completed, but results have not yet been published. Phase IIa trials are currently ongoing to assess the safety, tolerability, and immunogenicity of AEC/BC02 in patients aged 18 years and older with LTBI.121
Current recombinant live vaccines in clinical trials include VPM1002, MVA85A, ChAdOx185A, Ad5Ag85A, and TB/Flu-04L. VPM1002 was shown to have good safety and immunogenicity in healthy infants and adults in phase I and phase IIa clinical trials,122 and the results of phase IIb clinical trials showed that it was safer than BCG for the prevention of human immunodeficiency virus (HIV) exposed and unexposed infants.123 Phase III clinical trials evaluating VPM1002 are ongoing. Early trials of MVA85A demonstrated good safety and immunogenicity,124 but Phase IIb trial results indicated a lack of evidence for vaccine efficacy.125 Subsequent studies combined MVA85A with the simian adenovirus vaccine ChAdOx185A, revealing that both vaccines express the antigen Ag85A.126 In a Phase I clinical trial of ChAdOx185A administered intramuscularly in adults previously vaccinated with BCG, the vaccine was used either alone or as part of a boost strategy with MVA85A.127 Phase I clinical trials of Ad5Ag85A demonstrated good tolerability and immunogenicity in both BCG-vaccinated and non-vaccinated subjects, with stronger immune responses observed in volunteers previously vaccinated with BCG.128 A Phase I clinical trial conducted in Kazakhstan in BCG-vaccinated, Mtb T-cell-negative healthy adults has validated the safety and immunogenicity of TB/Flu-04L.129 Additionally, a Phase IIa clinical trial targeting LTBI populations is currently underway.130
In summary, following Mtb infection, the timely activation of both innate and adaptive immunity and the regulation of the balance between various immune cells and cytokines are crucial. Mtb is adept at disrupting the communication between innate and adaptive immunity. Understanding this mechanism is crucial for developing more effective vaccines and provides a theoretical foundation and direction for tuberculosis treatment.
Data Sharing Statement
No datasets were generated or analysed during the current study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The study was supported by National Natural Science Foundation of China (No. 82072460); National Natural Science Foundation of China (No. 82472469); Natural Science Foundation of Hunan Province (No. 2023JJ30878); Science and Technology Fund Project of Guizhou Provincial Health Commission (gzwkj2022-366).
Disclosure
All authors declare that they have no competing interests in this work.
References
1. World Health Organization. Global tuberculosis report 2024. 2024. Available from: https://wwwwhoint/news-room/fact-sheets/detail/tuberculosis.
2. Urbanowski ME, Ordonez AA, Ruiz-Bedoya CA, Jain SK, Bishai WR. Cavitary tuberculosis: the gateway of disease transmission. Lancet Infect Dis. 2020;20(6):e117–e128. doi:10.1016/s1473-3099(20)30148-1
3. Cohen A, Mathiasen VD, Schön T, Wejse C. The global prevalence of latent tuberculosis: a systematic review and meta-analysis. Eur Respir J. 2019;54(3):1900655. doi:10.1183/13993003.00655-2019
4. Sankar P, Mishra BB. Early innate cell interactions with Mycobacterium tuberculosis in protection and pathology of tuberculosis. Front Immunol. 2023;14:1260859. doi:10.3389/fimmu.2023.1260859
5. Rendón Ramírez EJ, Rosas-Taraco AG, Soto-Monciváis B, et al. Comparison of CD4+/CD8+ lymphocytic subpopulations pre- and post-antituberculosis treatment in patients with diabetes and tuberculosis. Pathogens. 2023;12(9):1181. doi:10.3390/pathogens12091181
6. Flynn JL, Chan J. Immune cell interactions in tuberculosis. Cell. 2022;185(25):4682–4702. doi:10.1016/j.cell.2022.10.025
7. Gupta T, Somanna N, Rowe T, et al. Ferrets as a model for tuberculosis transmission. Front Cell Infect Microbiol. 2022;12:873416. doi:10.3389/fcimb.2022.873416
8. Moreira-Teixeira L, Tabone O, Graham CM, et al. Mouse transcriptome reveals potential signatures of protection and pathogenesis in human tuberculosis. Nat Immunol. 2020;21(4):464–476. doi:10.1038/s41590-020-0610-z
9. Zhan L, Tang J, Sun M, Qin C. Animal models for tuberculosis in translational and precision medicine. Front Microbiol. 2017;8:717. doi:10.3389/fmicb.2017.00717
10. Kumar R, Kolloli A, Singh P, Vinnard C, Kaplan G, Subbian S. Thalidomide and phosphodiesterase 4 inhibitors as host directed therapeutics for tuberculous meningitis: insights from the rabbit model. Front Cell Infect Microbiol. 2019;9:450. doi:10.3389/fcimb.2019.00450
11. Hunter RL, Actor JK, Hwang SA, et al. Pathogenesis and animal models of post-primary (Bronchogenic) tuberculosis, a review. Pathogens. 2018;7(1):19. doi:10.3390/pathogens7010019
12. Luo G, Zeng D, Liu J, et al. Temporal and cellular analysis of granuloma development in mycobacterial infected adult zebrafish. J Leukoc Biol. 2024;115(3):525–535. doi:10.1093/jleuko/qiad145
13. Singh AK, Gupta UD. Animal models of tuberculosis: lesson learnt. Indian J Med Res. 2018;147(5):456–463. doi:10.4103/ijmr.IJMR_554_18
14. Liu CH, Liu H, Ge B. Innate immunity in tuberculosis: host defense vs pathogen evasion. Cell Mol Immunol. 2017;14(12):963–975. doi:10.1038/cmi.2017.88
15. Schneider BE, Behrends J, Hagens K, Harmel N, Shayman JA, Schaible UE. Lysosomal phospholipase A2: a novel player in host immunity to Mycobacterium tuberculosis. Eur J Immunol. 2014;44(8):2394–2404. doi:10.1002/eji.201344383
16. Weiss G, Schaible UE. Macrophage defense mechanisms against intracellular bacteria. Immunol Rev. 2015;264(1):182–203. doi:10.1111/imr.12266
17. Mihret A. The role of dendritic cells in Mycobacterium tuberculosis infection. Virulence. 2012;3(7):654–659. doi:10.4161/viru.22586
18. Peng X, Remacle JE, Kasran A, Huylebroeck D, Ceuppens JL. IL-12 up-regulates CD40 ligand (CD154) expression on human T cells. J Immunol. 1998;160(3):1166–1172.
19. Levin N, Pato A, Cafri G, et al. Spontaneous activation of antigen-presenting cells by genes encoding truncated homo-oligomerizing derivatives of CD40. J Immunother. 2017;40(2):39–50. doi:10.1097/cji.0000000000000150
20. Hilda JN, Das S, Tripathy SP, Hanna LE. Role of neutrophils in tuberculosis: a bird’s eye view. Innate Immun. 2020;26(4):240–247. doi:10.1177/1753425919881176
21. Braian C, Hogea V, Stendahl O. Mycobacterium tuberculosis- induced neutrophil extracellular traps activate human macrophages. J Innate Immun. 2013;5(6):591–602. doi:10.1159/000348676
22. Sampath P, Moideen K, Ranganathan UD, Bethunaickan R. Monocyte subsets: phenotypes and function in tuberculosis infection. Front Immunol. 2018;9:1726. doi:10.3389/fimmu.2018.01726
23. Sköld M, Behar SM. Tuberculosis triggers a tissue-dependent program of differentiation and acquisition of effector functions by circulating monocytes. J Immunol. 2008;181(9):6349–6360. doi:10.4049/jimmunol.181.9.6349
24. Castaño D, García LF, Rojas M. Increased frequency and cell death of CD16+ monocytes with Mycobacterium tuberculosis infection. Tuberculosis. 2011;91(5):348–360. doi:10.1016/j.tube.2011.04.002
25. Sánchez MD, García Y, Montes C, et al. Functional and phenotypic changes in monocytes from patients with tuberculosis are reversed with treatment. Microbes Infect. 2006;8(9–10):2492–2500. doi:10.1016/j.micinf.2006.06.005
26. Antonelli LR, Gigliotti Rothfuchs A, Gonçalves R, et al. Intranasal Poly-IC treatment exacerbates tuberculosis in mice through the pulmonary recruitment of a pathogen-permissive monocyte/macrophage population. J Clin Invest. 2010;120(5):1674–1682. doi:10.1172/jci40817
27. Samstein M, Schreiber HA, Leiner IM, Susac B, Glickman MS, Pamer EG. Essential yet limited role for CCR2+ inflammatory monocytes during Mycobacterium tuberculosis-specific T cell priming. Elife. 2013;2:e01086. doi:10.7554/eLife.01086
28. Vankayalapati R, Garg A, Porgador A, et al. Role of NK cell-activating receptors and their ligands in the lysis of mononuclear phagocytes infected with an intracellular bacterium. J Immunol. 2005;175(7):4611–4617. doi:10.4049/jimmunol.175.7.4611
29. Dhiman R, Indramohan M, Barnes PF, et al. IL-22 produced by human NK cells inhibits growth of Mycobacterium tuberculosis by enhancing phagolysosomal fusion. J Immunol. 2009;183(10):6639–6645. doi:10.4049/jimmunol.0902587
30. Schierloh P, Alemán M, Yokobori N, et al. NK cell activity in tuberculosis is associated with impaired CD11a and ICAM-1 expression: a regulatory role of monocytes in NK activation. Immunology. 2005;116(4):541–552. doi:10.1111/j.1365-2567.2005.02259.x
31. Paidipally P, Tripathi D, Van A, et al. Interleukin-21 regulates natural killer cell responses during Mycobacterium tuberculosis infection. J Infect Dis. 2018;217(8):1323–1333. doi:10.1093/infdis/jiy034
32. Kim H, Shin SJ. Pathological and protective roles of dendritic cells in Mycobacterium tuberculosis infection: interaction between host immune responses and pathogen evasion. Front Cell Infect Microbiol. 2022;12:891878. doi:10.3389/fcimb.2022.891878
33. Chen K, Kolls JK. T cell-mediated host immune defenses in the lung. Annu Rev Immunol. 2013;31:605–633. doi:10.1146/annurev-immunol-032712-100019
34. Gopal R, Monin L, Slight S, et al. Unexpected role for IL-17 in protective immunity against hypervirulent Mycobacterium tuberculosis HN878 infection. PLoS Pathog. 2014;10(5):e1004099. doi:10.1371/journal.ppat.1004099
35. Lyadova IV, Panteleev AV. Th1 and Th17 cells in tuberculosis: protection, pathology, and biomarkers. Mediators Inflamm. 2015;2015:854507. doi:10.1155/2015/854507
36. Jahandideh B, Derakhshani M, Abbaszadeh H, et al. The pro-Inflammatory cytokines effects on mobilization, self-renewal and differentiation of hematopoietic stem cells. Hum Immunol. 2020;81(5):206–217. doi:10.1016/j.humimm.2020.01.004
37. Yandrapally S, Agarwal A, Chatterjee A, Sarkar S, Mohareer K, Banerjee S. Mycobacterium tuberculosis EspR modulates Th1-Th2 shift by transcriptionally regulating IL-4, steering increased mycobacterial persistence and HIV propagation during co-infection. Front Immunol. 2023;14:1276817. doi:10.3389/fimmu.2023.1276817
38. Safar HA, Mustafa AS, Amoudy HA, El-Hashim A. The effect of adjuvants and delivery systems on Th1, Th2, Th17 and Treg cytokine responses in mice immunized with Mycobacterium tuberculosis-specific proteins. PLoS One. 2020;15(2):e0228381. doi:10.1371/journal.pone.0228381
39. Mills KHG. IL-17 and IL-17-producing cells in protection versus pathology. Nat Rev Immunol. 2023;23(1):38–54. doi:10.1038/s41577-022-00746-9
40. Nathan A, Beynor JI, Baglaenko Y, et al. Multimodally profiling memory T cells from a tuberculosis cohort identifies cell state associations with demographics, environment and disease. Nat Immunol. 2021;22(6):781–793. doi:10.1038/s41590-021-00933-1
41. Wang L, Ma H, Wen Z, et al. Single-cell RNA-sequencing reveals heterogeneity and intercellular crosstalk in human tuberculosis lung. J Infect. 2023;87(5):373–384. doi:10.1016/j.jinf.2023.09.004
42. Lin PL, Flynn JL. CD8 T cells and Mycobacterium tuberculosis infection. Semin Immunopathol. 2015;37(3):239–249. doi:10.1007/s00281-015-0490-8
43. Lees JR. CD8+ T cells: the past and future of immune regulation. Cell Immunol. 2020;357:104212. doi:10.1016/j.cellimm.2020.104212
44. Bénard A, Sakwa I, Schierloh P, et al. B cells producing type I IFN modulate macrophage polarization in tuberculosis. Am J Respir Crit Care Med. 2018;197(6):801–813. doi:10.1164/rccm.201707-1475OC
45. Kinker GS, Vitiello GAF, Ferreira WAS, Chaves AS, Cordeiro de Lima VC, Medina TDS. B cell orchestration of anti-tumor immune responses: a matter of cell localization and communication. Front Cell Dev Biol. 2021;9:678127. doi:10.3389/fcell.2021.678127
46. Sanaei MJ, Nahid-Samiei M, Abadi MSS, et al. New insights into regulatory B cells biology in viral, bacterial, and parasitic infections. Infect Genet Evol. 2021;89:104753. doi:10.1016/j.meegid.2021.104753
47. Ponnusamy N, Arumugam M. Interaction of host Pattern Recognition Receptors (PRRs) with mycobacterium tuberculosis and ayurvedic management of tuberculosis: a systemic approach. Infect Disord Drug Targets. 2022;22(2):e130921196420. doi:10.2174/1871526521666210913110834
48. Varshney D, Singh S, Sinha E, et al. Systematic review and meta-analysis of human Toll-like receptors genetic polymorphisms for susceptibility to tuberculosis infection. Cytokine. 2022;152:155791. doi:10.1016/j.cyto.2021.155791
49. Scanga CA, Bafica A, Feng CG, Cheever AW, Hieny S, Sher A. MyD88-deficient mice display a profound loss in resistance to Mycobacterium tuberculosis associated with partially impaired Th1 cytokine and nitric oxide synthase 2 expression. Infect Immun. 2004;72(4):2400–2404. doi:10.1128/iai.72.4.2400-2404.2004
50. Fremond CM, Yeremeev V, Nicolle DM, Jacobs M, Quesniaux VF, Ryffel B. Fatal Mycobacterium tuberculosis infection despite adaptive immune response in the absence of MyD88. J Clin Invest. 2004;114(12):1790–1799. doi:10.1172/jci21027
51. Tang X, Hu F, Xia X, et al. Mannose-capped lipoarabinomannan (ManLAM) binding TLR2 activates mast cells to release exosomes and induces M2 polarization of macrophages. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 2021;37(6):481–486.
52. Cai LX, Lei YY, Xie TY, et al. Overexpression of EspL inhibits autophagy and antigen presentation to promote the intracellular survival of Mycobacterium tuberculosis avirulent strains. Anim Dis. 2024;4(1):22. doi:10.1186/s44149-024-00128-9
53. Sengupta S, Nayak B, Meuli M, Sander P, Mishra S, Sonawane A. Mycobacterium tuberculosis phosphoribosyltransferase promotes bacterial survival in macrophages by inducing histone hypermethylation in autophagy-related genes. Front Cell Infect Microbiol. 2021;11:676456. doi:10.3389/fcimb.2021.676456
54. Feng S, McNehlan ME, Kinsella RL, et al. Autophagy promotes efficient T cell responses to restrict high-dose Mycobacterium tuberculosis infection in mice. Nat Microbiol. 2024;9(3):684–697. doi:10.1038/s41564-024-01608-x
55. Golovkine GR, Roberts AW, Morrison HM, et al. Autophagy restricts Mycobacterium tuberculosis during acute infection in mice. Nat Microbiol. 2023;8(5):819–832. doi:10.1038/s41564-023-01354-6
56. Zhai W, Wu F, Zhang Y, Fu Y, Liu Z. The immune escape mechanisms of Mycobacterium Tuberculosis. Int J Mol Sci. 2019;20(2):340. doi:10.3390/ijms20020340
57. Wong D, Chao JD, Av-Gay Y. Mycobacterium tuberculosis-secreted phosphatases: from pathogenesis to targets for TB drug development. Trends Microbiol. 2013;21(2):100–109. doi:10.1016/j.tim.2012.09.002
58. Ramon-Luing LA, Palacios Y, Ruiz A, Téllez-Navarrete NA, Chavez-Galan L. Virulence factors of Mycobacterium tuberculosis as modulators of cell death mechanisms. Pathogens. 2023;12(6):839. doi:10.3390/pathogens12060839
59. Auguste P, Tsertsvadze A, Pink J, et al. Comparing interferon-gamma release assays with tuberculin skin test for identifying latent tuberculosis infection that progresses to active tuberculosis: systematic review and meta-analysis. BMC Infect Dis. 2017;17(1):200. doi:10.1186/s12879-017-2301-4
60. Singh R, Dwivedi SP, Gaharwar US, Meena R, Rajamani P, Prasad T. Recent updates on drug resistance in Mycobacterium tuberculosis. J Appl Microbiol. 2020;128(6):1547–1567. doi:10.1111/jam.14478
61. Mancuso G, Midiri A, De Gaetano S, Ponzo E, Biondo C. Tackling drug-resistant tuberculosis: new challenges from the old pathogen Mycobacterium tuberculosis. Microorganisms. 2023;11(9):2277. doi:10.3390/microorganisms11092277
62. Kolonko M, Geffken AC, Blumer T, Hagens K, Schaible UE, Hagedorn M. WASH-driven actin polymerization is required for efficient mycobacterial phagosome maturation arrest. Cell Microbiol. 2014;16(2):232–246. doi:10.1111/cmi.12217
63. Bussi C, Gutierrez MG. Mycobacterium tuberculosis infection of host cells in space and time. FEMS Microbiol Rev. 2019;43(4):341–361. doi:10.1093/femsre/fuz006
64. Queval CJ, Song OR, Carralot JP, et al. Mycobacterium tuberculosis controls phagosomal acidification by targeting CISH-mediated signaling. Cell Rep. 2017;20(13):3188–3198. doi:10.1016/j.celrep.2017.08.101
65. Egen JG, Rothfuchs AG, Feng CG, Winter N, Sher A, Germain RN. Macrophage and T cell dynamics during the development and disintegration of mycobacterial granulomas. Immunity. 2008;28(2):271–284. doi:10.1016/j.immuni.2007.12.010
66. Clay H, Volkman HE, Ramakrishnan L. Tumor necrosis factor signaling mediates resistance to mycobacteria by inhibiting bacterial growth and macrophage death. Immunity. 2008;29(2):283–294. doi:10.1016/j.immuni.2008.06.011
67. Roca FJ, Whitworth LJ, Redmond S, Jones AA, Ramakrishnan L. TNF induces pathogenic programmed macrophage necrosis in tuberculosis through a mitochondrial-lysosomal-endoplasmic reticulum circuit. Cell. 2019;178(6):1344–1361.e11. doi:10.1016/j.cell.2019.08.004
68. Bourigault ML, Segueni N, Rose S, et al. Relative contribution of IL-1α, IL-1β and TNF to the host response to Mycobacterium tuberculosis and attenuated M. bovis BCG. Immun Inflamm Dis. 2013;1(1):47–62. doi:10.1002/iid3.9
69. Lee AM, Nathan CF. Type I interferon exacerbates Mycobacterium tuberculosis induced human macrophage death. EMBO Rep. 2024;25(7):3064–3089. doi:10.1038/s44319-024-00171-0
70. Zhang L, Jiang X, Pfau D, Ling Y, Nathan CF. Type I interferon signaling mediates Mycobacterium tuberculosis-induced macrophage death. J Exp Med. 2021;218(2):e20200887. doi:10.1084/jem.20200887
71. Saqib M, Das S, Nafiz TN, et al. Pathogenic role for CD101-negative neutrophils in the type I interferon-mediated immunopathogenesis of tuberculosis. Cell Rep. 2025;44(1):115072. doi:10.1016/j.celrep.2024.115072
72. Naik SK, McNehlan ME, Mreyoud Y, et al. Type I IFN signaling in the absence of IRGM1 promotes M. tuberculosis replication in immune cells by suppressing T cell responses. Mucosal Immunol. 2024;17(5):1114–1127. doi:10.1016/j.mucimm.2024.07.002
73. Kaufmann E, Sanz J, Dunn JL, et al. BCG educates hematopoietic stem cells to generate protective innate immunity against tuberculosis. Cell. 2018;172(1–2):176–190.e19. doi:10.1016/j.cell.2017.12.031
74. Chen J, Gao L, Wu X, et al. BCG-induced trained immunity: history, mechanisms and potential applications. J Transl Med. 2023;21(1):106. doi:10.1186/s12967-023-03944-8
75. Chai Q, Lu Z, Liu CH. Host defense mechanisms against Mycobacterium tuberculosis. Cell Mol Life Sci. 2020;77(10):1859–1878. doi:10.1007/s00018-019-03353-5
76. Ramos-Kichik V, Mondragón-Flores R, Mondragón-Castelán M, et al. Neutrophil extracellular traps are induced by Mycobacterium tuberculosis. Tuberculosis. 2009;89(1):29–37. doi:10.1016/j.tube.2008.09.009
77. Moreira-Teixeira L, Stimpson PJ, Stavropoulos E, et al. Type I IFN exacerbates disease in tuberculosis-susceptible mice by inducing neutrophil-mediated lung inflammation and NETosis. Nat Commun. 2020;11(1):5566. doi:10.1038/s41467-020-19412-6
78. Biron BM, Chung CS, O’Brien XM, Chen Y, Reichner JS, Ayala A. Cl-amidine prevents histone 3 citrullination and neutrophil extracellular trap formation, and improves survival in a murine sepsis model. J Innate Immun. 2017;9(1):22–32. doi:10.1159/000448808
79. Thålin C, Daleskog M, Göransson SP, et al. Validation of an enzyme-linked immunosorbent assay for the quantification of citrullinated histone H3 as a marker for neutrophil extracellular traps in human plasma. Immunol Res. 2017;65(3):706–712. doi:10.1007/s12026-017-8905-3
80. Kuczia P, Zuk J, Iwaniec T, et al. Citrullinated histone H3, a marker of extracellular trap formation, is increased in blood of stable asthma patients. Clin Transl Allergy. 2020;10:31. doi:10.1186/s13601-020-00337-8
81. Nwongbouwoh Muefong C, Owolabi O, Donkor S, et al. Neutrophils contribute to severity of tuberculosis pathology and recovery from lung damage pre- and posttreatment. Clin Infect Dis. 2022;74(10):1757–1766. doi:10.1093/cid/ciab729
82. Keller C, Hoffmann R, Lang R, Brandau S, Hermann C, Ehlers S. Genetically determined susceptibility to tuberculosis in mice causally involves accelerated and enhanced recruitment of granulocytes. Infect Immun. 2006;74(7):4295–4309. doi:10.1128/iai.00057-06
83. Pedrosa J, Saunders BM, Appelberg R, Orme IM, Silva MT, Cooper AM. Neutrophils play a protective nonphagocytic role in systemic Mycobacterium tuberculosis infection of mice. Infect Immun. 2000;68(2):577–583. doi:10.1128/iai.68.2.577-583.2000
84. Balboa L, Romero MM, Basile JI, et al. Paradoxical role of CD16+CCR2+CCR5+ monocytes in tuberculosis: efficient APC in pleural effusion but also mark disease severity in blood. J Leukoc Biol. 2011;90(1):69–75. doi:10.1189/jlb.1010577
85. Chu J, Gao F, Yan M, et al. Natural killer cells: a promising immunotherapy for cancer. J Transl Med. 2022;20(1):240. doi:10.1186/s12967-022-03437-0
86. Esin S, Counoupas C, Aulicino A, et al. Interaction of Mycobacterium tuberculosis cell wall components with the human natural killer cell receptors NKp44 and Toll-like receptor 2. Scand J Immunol. 2013;77(6):460–469. doi:10.1111/sji.12052
87. Venkatasubramanian S, Cheekatla S, Paidipally P, et al. IL-21-dependent expansion of memory-like NK cells enhances protective immune responses against Mycobacterium tuberculosis. Mucosal Immunol. 2017;10(4):1031–1042. doi:10.1038/mi.2016.105
88. Choreño-Parra JA, Jiménez-álvarez LA, Muñoz-Torrico M, et al. Antigens of Mycobacterium tuberculosis stimulate CXCR6+ natural killer cells. Front Immunol. 2020;11:582414. doi:10.3389/fimmu.2020.582414
89. Choreño Parra JA, Martínez Zúñiga N, Jiménez Zamudio LA, Jiménez Álvarez LA, Salinas Lara C, Zúñiga J. Memory of Natural Killer Cells: a New Chance against Mycobacterium tuberculosis? Front Immunol. 2017;8:967. doi:10.3389/fimmu.2017.00967
90. Anderson DA, Dutertre CA, Ginhoux F, Murphy KM. Genetic models of human and mouse dendritic cell development and function. Nat Rev Immunol. 2021;21(2):101–115. doi:10.1038/s41577-020-00413-x
91. Srivastava S, Ernst JD. Cell-to-cell transfer of M. tuberculosis antigens optimizes CD4 T cell priming. Cell Host Microbe. 2014;15(6):741–752. doi:10.1016/j.chom.2014.05.007
92. Dong C. Cytokine Regulation and Function in T Cells. Annu Rev Immunol. 2021;39:51–76. doi:10.1146/annurev-immunol-061020-053702
93. Belyakov IM, Ahlers JD. Functional CD8+ CTLs in mucosal sites and HIV infection: moving forward toward a mucosal AIDS vaccine. Trends Immunol. 2008;29(11):574–585. doi:10.1016/j.it.2008.07.010
94. de Martino M, Lodi L, Galli L, Chiappini E. Immune response to Mycobacterium tuberculosis: a narrative review. Front Pediatr. 2019;7:350. doi:10.3389/fped.2019.00350
95. Guo L, Hu-Li J, Zhu J, et al. In TH2 cells the Il4 gene has a series of accessibility states associated with distinctive probabilities of IL-4 production. Proc Natl Acad Sci U S A. 2002;99(16):10623–10628. doi:10.1073/pnas.162360199
96. Woodworth JS, Wu Y, Behar SM. Mycobacterium tuberculosis-specific CD8+ T cells require perforin to kill target cells and provide protection in vivo. J Immunol. 2008;181(12):8595–8603. doi:10.4049/jimmunol.181.12.8595
97. Noschka R, Wondany F, Kizilsavas G, et al. Gran1: a granulysin-derived peptide with potent activity against intracellular Mycobacterium tuberculosis. Int J Mol Sci. 2021;22(16):8392. doi:10.3390/ijms22168392
98. Yang JD, Mott D, Sutiwisesak R, et al. Mycobacterium tuberculosis-specific CD4+ and CD8+ T cells differ in their capacity to recognize infected macrophages. PLoS Pathog. 2018;14(5):e1007060. doi:10.1371/journal.ppat.1007060
99. Naderi W, Schreiner D, King CG. T-cell-B-cell collaboration in the lung. Curr Opin Immunol. 2023;81:102284. doi:10.1016/j.coi.2023.102284
100. Abebe F, Belay M, Legesse M, KLMC F, Ottenhoff THM. IgA and IgG against Mycobacterium tuberculosis Rv2031 discriminate between pulmonary tuberculosis patients, Mycobacterium tuberculosis-infected and non-infected individuals. PLoS One. 2018;13(1):e0190989. doi:10.1371/journal.pone.0190989
101. Rodrigues LC, Diwan VK, Wheeler JG. Protective effect of BCG against tuberculous meningitis and miliary tuberculosis: a meta-analysis. Int J Epidemiol. 1993;22(6):1154–1158. doi:10.1093/ije/22.6.1154
102. Khader SA, Divangahi M, Hanekom W, et al. Targeting innate immunity for tuberculosis vaccination. J Clin Invest. 2019;129(9):3482–3491. doi:10.1172/jci128877
103. Fuertes Marraco SA, Soneson C, Cagnon L, et al. Long-lasting stem cell-like memory CD8+ T cells with a naïve-like profile upon yellow fever vaccination. Sci Transl Med. 2015;7(282):282ra48. doi:10.1126/scitranslmed.aaa3700
104. Akondy RS, Fitch M, Edupuganti S, et al. Origin and differentiation of human memory CD8 T cells after vaccination. Nature. 2017;552(7685):362–367. doi:10.1038/nature24633
105. Baldridge MT, King KY, Boles NC, Weksberg DC, Goodell MA. Quiescent haematopoietic stem cells are activated by IFN-gamma in response to chronic infection. Nature. 2010;465(7299):793–797. doi:10.1038/nature09135
106. Schüller S, Neefjes J, Ottenhoff T, Thole J, Young D. Coronin is involved in uptake of Mycobacterium bovis BCG in human macrophages but not in phagosome maintenance. Cell Microbiol. 2001;3(12):785–793. doi:10.1046/j.1462-5822.2001.00155.x
107. Schrager LK, Vekemens J, Drager N, Lewinsohn DM, Olesen OF. The status of tuberculosis vaccine development. Lancet Infect Dis. 2020;20(3):e28–e37. doi:10.1016/s1473-3099(19)30625-5
108. Gonzalo-Asensio J, Marinova D, Martin C, Aguilo N. MTBVAC: attenuating the human pathogen of Tuberculosis (TB) toward a promising vaccine against the TB epidemic. Front Immunol. 2017;8:1803. doi:10.3389/fimmu.2017.01803
109. Tameris M, Mearns H, Penn-Nicholson A, et al. Live-attenuated Mycobacterium tuberculosis vaccine MTBVAC versus BCG in adults and neonates: a randomised controlled, double-blind dose-escalation trial. Lancet Respir Med. 2019;7(9):757–770. doi:10.1016/s2213-2600(19)30251-6
110. Martín C, Marinova D, Aguiló N, Gonzalo-Asensio J. MTBVAC, a live TB vaccine poised to initiate efficacy trials 100 years after BCG. Vaccine. 2021;39(50):7277–7285. doi:10.1016/j.vaccine.2021.06.049
111. Prabowo SA, Painter H, Zelmer A, et al. RUTI vaccination enhances inhibition of mycobacterial growth ex vivo and induces a shift of monocyte phenotype in mice. Front Immunol. 2019;10:894. doi:10.3389/fimmu.2019.00894
112. Nell AS, D’Lom E, Bouic P, et al. Safety, tolerability, and immunogenicity of the novel antituberculous vaccine RUTI: randomized, placebo-controlled phase II clinical trial in patients with latent tuberculosis infection. PLoS One. 2014;9(2):e89612. doi:10.1371/journal.pone.0089612
113. Sharma SK, Katoch K, Sarin R, et al. Efficacy and Safety of Mycobacterium indicus pranii as an adjunct therapy in Category II pulmonary tuberculosis in a randomized trial. Sci Rep. 2017;7(1):3354. doi:10.1038/s41598-017-03514-1
114. Masonou T, Hokey DA, Lahey T, et al. CD4+ T cell cytokine responses to the DAR-901 booster vaccine in BCG-primed adults: a randomized, placebo-controlled trial. PLoS One. 2019;14(5):e0217091. doi:10.1371/journal.pone.0217091
115. Weng H, Huang JY, Meng XY, Li S, Zhang GQ. Adjunctive therapy of Mycobacterium vaccae vaccine in the treatment of multidrug-resistant tuberculosis: a systematic review and meta-analysis. Biomed Rep. 2016;4(5):595–600. doi:10.3892/br.2016.624
116. Kumarasamy N, Poongulali S, Bollaerts A, et al. A randomized, controlled safety, and immunogenicity trial of the M72/AS01 candidate tuberculosis vaccine in HIV-positive Indian adults. Medicine. 2016;95(3):e2459. doi:10.1097/md.0000000000002459
117. Gates, wellcome rekindle TB vaccine. Nat Biotechnol. 2023;41(8):1039. doi:10.1038/s41587-023-01914-5
118. Suliman S, Luabeya AKK, Geldenhuys H, et al. Dose optimization of H56:IC31 vaccine for tuberculosis-endemic populations. A double-blind, placebo-controlled, dose-selection trial. Am J Respir Crit Care Med. 2019;199(2):220–231. doi:10.1164/rccm.201802-0366OC
119. Munseri P, Said J, Amour M, et al. DAR-901 vaccine for the prevention of infection with Mycobacterium tuberculosis among BCG-immunized adolescents in Tanzania: a randomized controlled, double-blind Phase 2b trial. Vaccine. 2020;38(46):7239–7245. doi:10.1016/j.vaccine.2020.09.055
120. Tkachuk AP, Bykonia EN, Popova LI, et al. Safety and Immunogenicity of the GamTBvac, the recombinant subunit tuberculosis vaccine candidate: a Phase II, multi-center, double-blind, randomized, placebo-controlled study. Vaccines. 2020;8(4):652. doi:10.3390/vaccines8040652
121. Guo X, Lu J, Li J, et al. The subunit AEC/BC02 vaccine combined with antibiotics provides protection in Mycobacterium tuberculosis-Infected Guinea Pigs. Vaccines. 2022;10(12):2164. doi:10.3390/vaccines10122164
122. Loxton AG, Knaul JK, Grode L, et al. Safety and immunogenicity of the recombinant mycobacterium bovis BCG vaccine VPM1002 in HIV-unexposed newborn infants in South Africa. Clin Vaccine Immunol. 2017;24(2):e00439–16. doi:10.1128/cvi.00439-16
123. Cotton MF, Madhi SA, Luabeya AK, et al. Safety and immunogenicity of VPM1002 versus BCG in South African newborn babies: a randomised, phase 2 non-inferiority double-blind controlled trial. Lancet Infect Dis. 2022;22(10):1472–1483. doi:10.1016/s1473-3099(22)00222-5
124. Scriba TJ, Tameris M, Mansoor N, et al. Dose-finding study of the novel tuberculosis vaccine, MVA85A, in healthy BCG-vaccinated infants. J Infect Dis. 2011;203(12):1832–1843. doi:10.1093/infdis/jir195
125. Ndiaye BP, Thienemann F, Ota M, et al. Safety, immunogenicity, and efficacy of the candidate tuberculosis vaccine MVA85A in healthy adults infected with HIV-1: a randomised, placebo-controlled, phase 2 trial. Lancet Respir Med. 2015;3(3):190–200. doi:10.1016/s2213-2600(15)00037-5
126. Hu Z, Lu SH, Lowrie DB, Fan XY. Research advances for virus-vectored tuberculosis vaccines and latest findings on tuberculosis vaccine development. Front Immunol. 2022;13:895020. doi:10.3389/fimmu.2022.895020
127. Wilkie M, Satti I, Minhinnick A, et al. A phase I trial evaluating the safety and immunogenicity of a candidate tuberculosis vaccination regimen, ChAdOx1 85A prime - MVA85A boost in healthy UK adults. Vaccine. 2020;38(4):779–789. doi:10.1016/j.vaccine.2019.10.102
128. Jeyanathan M, Damjanovic D, Yao Y, Bramson J, Smaill F, Xing Z. Induction of an immune-protective T-cell repertoire with diverse genetic coverage by a novel viral-vectored tuberculosis vaccine in humans. J Infect Dis. 2016;214(12):1996–2005. doi:10.1093/infdis/jiw467
129. Buzitskaya Z, Stosman K, Khairullin B, et al. A new intranasal influenza vector-based vaccine TB/FLU-04L against tuberculosis: preclinical safety studies. Drug Res. 2022;72(5):255–258. doi:10.1055/a-1785-3936
130. Li J, Zhao A, Tang J, et al. Tuberculosis vaccine development: from classic to clinical candidates. Eur J Clin Microbiol Infect Dis. 2020;39(8):1405–1425. doi:10.1007/s10096-020-03843-6
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.