")
Back to Journals » Drug Design, Development and Therapy » Volume 18
Impact of Lipids on Insulin Resistance: Insights from Human and Animal Studies
Authors Elkanawati RY, Sumiwi SA, Levita J
Received 10 March 2024
Accepted for publication 24 July 2024
Published 31 July 2024 Volume 2024:18 Pages 3337—3360
DOI https://doi.org/10.2147/DDDT.S468147
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tuo Deng
Rani Yulifah Elkanawati,1,* Sri Adi Sumiwi,2 Jutti Levita2,*
1Master Program in Pharmacy, Faculty of Pharmacy, Padjadjaran University, Jawa Barat, West Java, 45363, Indonesia; 2Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Padjadjaran University, Sumedang, West Java, 45363, Indonesia
*These authors contributed equally to this work
Correspondence: Jutti Levita, Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Padjadjaran University, Sumedang, West Java, 46363, Indonesia, Tel +6222-84288888 Ext 3510, Email [email protected]
Abstract: Insulin resistance (IR) is a complex pathological condition central to metabolic diseases such as type 2 diabetes mellitus (T2DM), cardiovascular disease, non-alcoholic fatty liver disease, and polycystic ovary syndrome (PCOS). This review evaluates the impact of lipids on insulin resistance (IR) by analyzing findings from human and animal studies. The articles were searched on the PubMed database using two keywords: (1) “Role of Lipids AND Insulin Resistance AND Humans” and (2) “Role of Lipids AND Insulin Resistance AND Animal Models”. Studies in humans revealed that elevated levels of free fatty acids (FFAs) and triglycerides (TGs) are closely associated with reduced insulin sensitivity, and interventions like metformin and omega-3 fatty acids show potential benefits. In animal models, high-fat diets disrupt insulin signaling and increase inflammation, with lipid mediators such as diacylglycerol (DAG) and ceramides playing significant roles. DAG activates protein kinase C, which eventually impairs insulin signaling, while ceramides inhibit Akt/PKB, further contributing to IR. Understanding these mechanisms is crucial for developing effective prevention and treatment strategies for IR-related diseases.
Keywords: high-fat diet, insulin resistance, lipid profile, type 2 diabetes mellitus
Introduction
Insulin resistance (IR) is a complex pathological condition that has become a focal point in studies related to metabolic diseases such as type 2 diabetes mellitus (T2DM), cardiovascular disease (CVD), non-alcoholic fatty liver disease (NAFLD), and polycystic ovary syndrome (PCOS).1 IR is characterized by the unresponsiveness of cells to insulin, which has a significant impact on global public health due to increasing obesity and unhealthy lifestyles.2,3 Lipids, such as diacylglycerol (DAG) and sphingolipids, are key mediators of impaired insulin sensitivity.4 DAG, for example, activates protein kinase C (PKC) which in turn inhibits the insulin signaling pathway through serine/threonine phosphorylation of insulin receptor substrate 1 and insulin receptor substrate 2 (IRS-1 and IRS-2).5 Sphingolipids such as ceramide cause stress on the endoplasmic reticulum (ER) and mitochondrial dysfunction, both of which contribute to IR. Ceramide inhibits the Akt signaling pathway by activating protein phosphatase 2A (PP2A), ultimately impairing glucose absorption and nutrient storage. In addition, the accumulation of ceramide in insulin-responsive tissues, such as muscle and liver, impairs insulin sensitivity and contributes to the development of obesity.6
As dietary patterns change, for example, the consumption of high-fat foods and carbohydrates increases, the necessity for an in-depth study of the mechanisms of lipid-IR interaction in metabolic diseases is crucial. Human studies, in the form of observational research and clinical trials, may provide important insights into the relationship between lipids and IR. Many studies in humans consistently show that increased levels of circulating free fatty acids (FFAs) and triglycerides (TGs) correlate with decreased insulin sensitivity,7–9 leading to fat accumulation in non-adipose tissues like muscle and liver, thus further inhibiting insulin action.10 Nevertheless, to understand the molecular mechanisms underlying this relationship, animal models have become a valuable research platform.
Animal models, such as rats and mice, offer deeper insights into the impact of lipids in IR. Dietary manipulations with high-fat diet (HFD) in these models reveal complex interactions between lipids and insulin pathways, proofing that lipids directly influence insulin sensitivity, accelerate hepatic lipid deposition, and eventually lead to IR in a relatively short time.11–15 Additionally, animal models allow researchers to investigate the long-term effects of lipid exposure on IR, which is difficult to do in human studies.
In this context, this review aims to evaluate the impact of lipids in causing IR, focusing on findings from studies in humans and animal models. The ways that lipids and IR are connected will be discussed, along with what these results mean for creating ways to prevent and treat conditions that are linked to IR. By bringing together findings on both models, it is anticipated that this review may provide deeper insights into the complexity of interactions between lipid and insulin pathways, as well as identify promising future research directions in this area.
Methods
To collect relevant information about the role of lipids in inducing IR in humans and animal models, a literature search was conducted in the PubMed database, limited to articles published from 2014 to 2024 to ensure that this review captures recent developments in understanding the impact of lipids in IR. The search was carried out using two keywords: (1) “Role of Lipids AND Insulin Resistance AND Humans”, which resulted in a total of 145 articles; (2) “Role of Lipids AND Insulin Resistance AND Animal Models”, which resulted in a total of 456 articles. Following the initial search, articles were screened based on title and abstract to exclude articles that were not directly relevant to the topic. The full texts of the remaining articles were retrieved and evaluated according to the inclusion criteria (free full-text articles written in English, studies related to lipid-induced IR, and studies directly related to the topic of the role of lipids in IR in humans and animal models). Finally, 62 articles met the criteria and were selected for review, as depicted in Figure 1. Additional searches were conducted as needed to ensure comprehensive coverage of the mechanisms underlying the relationship between lipids and IR.
 |
Figure 1 Literature review search method. |
Results
Of the 62 articles, 12 articles described studies in humans (Table 1), and 50 articles confirmed studies in animal models (Table 2).
 |
Table 1 Relationship Between Lipids and Insulin Resistance in Humans |
 |
Table 2 Relationship Between Lipids and Insulin Resistance in Animal Models |
The total participants and/or patients included in the 12 articles were 952 males (933 of those were with T2DM) and 799 females (597 of those were with T2DM). Only 6 of the 12 articles delineate ethnicity, with Chinese being the largest group in the studies. Studies that have been conducted regarding IR involve the participation of two main groups of patients, namely diabetic patients (5 studies), PCOS patients (2 studies), and others (5 studies). Of the five studies conducted on T2DM patients, various aspects related to the management of IR and changes in lipid metabolism were reported. However, the variations in ethnicity and gender diversity in human studies may contribute to the limitation of this review. Furthermore, various types of design studies used, such as RCT (randomized controlled trial), double-blind or single-blind, cross-over, and open-label or placebo-controlled, duration of the study, and the interventions used (synthetic drugs or plant-derived drugs), have also contributed to the limitations of this review.
Of the 50 articles on animal studies, only 3 included the use of extracts from plants as part of their research, including the extraction method and solvent used for extraction. In animal models, a high-fat diet is given to create an animal model of IR. The duration of HFD induction in the study ranged from 4 to 30 weeks. Various types of experimental animals were used, with various characteristics, such as varying ages and different genders, with the majority of male mice, and some were mice with special conditions such as diabetes or obesity. Certain demographic data, such as age, body weight, and sample size, are also included in the results of this review, which can provide information about the animal population used in the experiments. A total of 24 articles mentioned the use of C57BL/6J mice, which is the most widely used type of mouse. The C57BL/6J mouse strain is widely used in metabolic research as an animal model for diet-induced obesity (DIO).71
This study used standard care drugs; for example, 4 articles mentioned metformin (biguanide class, for treating T2DM),11,12,26,27 2 articles mentioned pioglitazone (thiazolidinedione class, for treating T2DM),28,29 and 1 article each mentioned amitriptyline (tricyclic antidepressant)30 and orlistat (weight loss medication).31 Based on the research carried out, the results show that exposure to lipids in humans and experimental animals can contribute to IR, and IR is the main trigger factor related to the development of T2DM.
Diabetes Mellitus (DM)
DM is prevalent worldwide and increasing annually. The International Diabetes Federation (IDF) ranks Indonesia 5th globally, after China, India, Pakistan, and America, for the highest number of adults (20–79 years) with DM in 2021 and 2045.72 According to WHO, DM is marked by increased blood glucose levels, known as hyperglycemia,73 occurring when the body cannot produce or effectively use insulin.74 Insulin, synthesized by pancreatic beta-cells, controls blood glucose levels, facilitates cell respiration, and is crucial for protein and fat metabolism.72
IR, the main trigger for T2DM, occurs when insulin cannot effectively stimulate glucose uptake or when target cells (muscle, adipose tissue, liver) respond poorly to insulin.75,76 This leads to hyperinsulinemia, and over time, pancreatic exhaustion results in elevated blood glucose and T2DM.77 IR also contributes to complications, such as heart disease, stroke, high blood pressure, and other metabolic problems.78–80
Obesity is a major risk factor for IR and T2DM.81,82 Obesity is the excessive accumulation of fat due to an imbalance in food intake and expenditure and can cause various health problems.77 The main cause of obesity is an unhealthy lifestyle and a poor diet, such as one high in fat and sugar, followed by a lack of physical activity.83,84 HFD can increase the number of small adipocytes, leading to fat accumulation in the liver and decreased insulin sensitivity.85 In obese individuals, there is increased production of FFAs, reactive oxygen species (ROS), and proinflammatory cytokines, which may contribute to the development of IR.86 An increase in FFAs concentration occurs for several reasons, such as greater FFA release due to increased adipose tissue mass, and FFA clearance may also be impaired.77 Consequently, high concentrations of FFA inhibit the antilipolytic activity of insulin and increase the rate of FFA release into the circulation.87 Therefore, it is important to adopt a healthy lifestyle, such as partaking in healthy foods, avoiding excessive calorie consumption, exercising regularly, and maintaining an ideal body weight.88,89 A vegan diet, which excludes animal products, may help in reducing body weight, fat mass, and visceral fat volume, as well as improve insulin sensitivity.21
Metformin is a commonly used antihyperglycemic agent in the management of T2DM, working by increasing insulin sensitivity and reducing glucose production by the liver.90 Omega-3 fatty acids, found in fatty fish, also play an important role in improving insulin function by increasing insulin sensitivity, reducing ceramide content, and increasing adiponectin expression in animal models,91 while reducing inflammation and IR.92
With a better understanding of the link between DM and IR, as well as appropriate prevention efforts, it is hoped that we can reduce the burden of this disease on individuals and society as a whole. Prevention and appropriate management can help reduce the risk of serious complications and improve the quality of life for those living with diabetes.
Studies in Humans
Studies in humans reveal that lipid accumulation, particularly saturated fats, triggers the occurrence of IR.21,22,93 These lipids, including FFAs and TGs, disrupt insulin signal transduction, produce pro-inflammatory cytokines, and oxidative stress, and inhibit intracellular signaling pathways.77 Ceramide is also a key mediator of IR.94 Understanding these lipids could lead to more effective prevention and treatment strategies for IR and related diseases.
Several studies highlight the impact of lipid infusions on glucose and insulin metabolism in the context of IR. Lipid infusions can significantly affect glucose and insulin metabolism, leading to IR. It increases the levels of TG and FFAs in the blood, worsening glucose tolerance and insulin sensitivity. Lipids affect pancreatic beta-cell function, causing insulin hypersecretion. However, decreased insulin clearance also contributes to hyperinsulinemia.23 Research has identified proteins that respond to intralipid and insulin infusions in healthy women and women with PCOS. Intralipid infusion reduces the ability of insulin to stimulate glucose utilization, indicating increased IR. In addition, intralipid infusion decreased fibroblast growth factor 19 (FGF19) levels, indicating impaired regulation of glucose and lipid metabolism.19
Lipid excess can lead to the accumulation of intramyocellular lipid metabolites (IMCL), which coincides with impaired insulin response. The fat composition in IMCL, especially saturated fat, has an important role in mediating IR.22 Palmitic acid, a saturated fat found in muscle TG, is associated with IR through increased concentrations of ceramide, which interferes with insulin activation of cell receptors and downstream signaling molecules such as IRS-1 and Akt/protein kinase B (PKB).95,96
Female sex hormones also play a role in protecting against lipid-mediated IR. Postmenopausal women, who naturally have increased levels of saturated fat, tend to accumulate more ceramides in response to high lipid loads, thereby increasing the risk of IR. This suggests that premenopausal women’s hormones may have a protective effect against ceramide accumulation and associated IR.24,32 Understanding the lipid composition in IMCL and the influence of female sex hormones may provide new insights into the management of IR and diabetes risk.
Ceramide, a type of fat in cells, has been linked to lipid-mediated skeletal muscle IR.97–99 Research has shown that individuals with T2DM have higher blood levels of ceramide,100 which is associated with IR, insulin sensitivity, and cardiovascular risk.101 This is consistent with a previous study showing obese and diabetic patients have higher levels of ceramide in their muscles.102 These findings support the role of ceramide in skeletal muscle IR.
Chiglitazar and lobeglitazone show potential for managing T2DM. Chiglitazar, a PPAR agonist (peroxisome proliferator-activated receptor), activates all three PPAR types (-alpha, -gamma, -delta), controlling glucose and fat levels, decreasing TG, increasing HDL-cholesterol, and enhancing insulin sensitivity.7,8 Lobeglitazone, a PPAR-gamma agonist, improves insulin sensitivity, reducing IR. Both drugs improve glycemic control, lipid regulation, and IR in T2DM patients, reducing disease complications.16
Natural substances can improve IR in T2DM and PCOS. Berberine, an alkaloid in Chinese herbal medicine, increases glucose consumption and absorption in cells, inhibits alpha-glucosidase, and improves insulin sensitivity.25 Cranberries, rich in polyphenols such as proanthocyanidins, inhibit carbohydrate digestive enzymes, decrease intestinal glucose absorption, and modulate inflammatory biomarkers, all of which contribute to improved insulin sensitivity.17 Anthocyanins in colored fruits and vegetables improve insulin sensitivity by increasing adiponectin expression and fatty acid oxidation.18 Overall, these three natural substances have a complementary role in improving IR, demonstrating their potential as therapeutic agents in managing T2DM and related metabolic conditions.
Studies in Animal Models
Various studies show that mice treated with an HFD experience increased glucose intolerance and IR,37 characterized by increased fasting blood glucose (FBG), insulin,51 total cholesterol, TG,68 and FFA levels, as well as decreased adiponectin levels.53 The underlying mechanisms involve complex biological pathways.
One of the main mechanisms is changes in insulin signaling in the liver and skeletal muscle. Signaling cascades such as IRS-1/Akt/GSK-3β (Glycogen Synthase Kinase-3 beta) have a major role in the regulation of glucose transport, glycogen synthesis, and energy metabolism.103 Increased Ser307 phosphorylation of IRS-1 characterizes peripheral IR, inhibits Glucose Transporter Type 4 (GLUT4) translocation to the cell surface, and causes hyperglycemia.104
In addition, HFD elevates the concentration of sphingolipids, such as ceramide, which interfere with the insulin signaling pathway by inhibiting insulin-induced phosphorylation of Akt/PKB in skeletal muscle and liver. This inhibition of Akt/PKB leads to the inhibition of GSK-3 α/β phosphorylation at Ser21/Ser9, resulting in impaired insulin sensitivity.13,15
HFD also triggers low-grade inflammation in adipose tissue, which in turn leads to the recruitment of immune cells and the production of proinflammatory cytokines, leading to IR.55 Studies in mice show that increased plasma FFA could activate muscle NF-kappaB signaling,105 which eventually leads to higher levels of proinflammatory cytokines such as tumor necrosis factor-alpha (TNF-α), interleukin-1 beta (IL-1β), IL-6, and Monocyte Chemoattractant Protein-1 (MCP-1).106 This increase in plasma MCP-1 can promote the migration of monocytes from the blood to adipose tissue. When macrophages settle in adipose tissue, they differentiate into inflammatory macrophages, which can release a large number of inflammatory factors.107 Additionally, HFD-induced increases in FFA also increase Toll Like Receptor-4 (TLR-4) activity, which may trigger inflammatory signaling pathways that contribute to the development of IR and atherosclerosis.68
Overall, this review shows that HFD significantly escalates the risk of developing IR in mice. The multiple mechanisms involved highlight the complexity of the role of HFDs in the development of IR, which has significant implications for metabolic health and the development of diseases such as T2DM. Therefore, a better understanding of these mechanisms is important for the development of appropriate prevention and treatment strategies.
This review discusses the unique role of proteins such as TNFSF14 (Tumor Necrosis Factor Superfamily Member 14) and NGBR (Nogo‐B Receptor) in influencing glucose metabolism and IR. TNFSF14, specifically Peptide 7, increases insulin signaling and fatty acid oxidation in skeletal muscle cells, improving glucose management and IR in mice fed with HFD.37 Overexpression of NGBR suggests that this protein functions in the improvement of insulin sensitivity and the reduction of endoplasmic reticulum stress in the liver and skeletal muscle.38
Bioactive compounds have also been identified as potential treatments for DM. Polygonatum sibiricum saponin (PSS) has demonstrated its ability to reduce IR, improve pancreatic beta-cell function, and facilitate the normal biological role of insulin. The mechanism of action of PSS involves increasing Akt which triggers GLUT4, thereby helping control blood glucose and improving glucose tolerance.26 Berberine, a bioactive compound from Rhizoma coptidis, shows potential in improving IR by reducing the accumulation of ceramide, which interferes with insulin signaling and activates AMP-activated protein kinase (AMPK) signaling.13 Cannabidiol (CBD) from Cannabis sativa lessens the accumulation of sphingolipids and exhibits anti-inflammatory and antioxidant effects that help improve the condition of IR and other diabetic complications.15 Stevioside, found in Stevia rebaudiana leaves, increases glucose uptake and oxidation in muscles by activating the IR/IRS-1/Akt/GLUT4 pathway and inhibiting oxidative stress.12 Not only that, resveratrol and cucurbitacin E also indicate the potential to overcome IR. Resveratrol increases glucose uptake via the PI3K/Akt signaling pathway to attenuate IR,11 while cucurbitacin E affects several pathways, including the inhibition of Janus Kinase/Signal Transducers and Activators of Transcription (JAK-STAT5), influences adipokines, decreases lipogenesis, activates AMPK, and regulates insulin signaling.31
Glucagon-like Peptide-1 (GLP-1) analogs, synthetic versions of the GLP-1 hormone, regulate glucose levels with a prolonged effect.108,109 They increase insulin release, reduce glucagon release, and slow gastric emptying.110 GLP-1 analogs maintain healthy blood glucose by modulating calcium signals in hepatocytes and reducing lipid accumulation.40 They restore insulin sensitivity by promoting GLUT4 translocation, facilitating glucose uptake, and reducing blood glucose. In HFD-fed mice, GLP-1 restored reduced phosphoinositide 3-kinase (PI3K) expression, improving the insulin signaling pathway.58
High levels of pancreastatin can cause IR in T2DM. PST inhibitor peptide-8 (PSTi8) improves insulin sensitivity in mice with IR by increasing glucose clearance, glycogenesis, glycolysis, and gluconeogenesis. PSTi8 targets the IRS1/2-PI3K-Akt signaling pathway, making it a promising therapeutic agent for metabolic diseases, particularly DM.80
The intricate interplay of lipid accumulation, inflammatory responses, and disrupted insulin signaling pathways underscores the multifaceted mechanisms through which an HFD-induced IR, highlighting the critical need for targeted prevention and therapeutic strategies to mitigate metabolic diseases such as T2DM.
Lipid-Induced Insulin Resistance
Adipose tissue, composed of adipocytes, can expand and store lipids through hypertrophy (enlargement of the cell size of the adipocyte) or hyperplasia (proliferation). When fat absorption increases, hypertrophic adipocytes stimulate hyperplastic adipocytes, which then attract more macrophages to enter and release inflammatory substances called adipokines. Over time, these hypertrophic adipocytes become dysfunctional and highly lipolytic, producing excessive FFA and contributing to the development of IR. This process is facilitated by the elastic properties of the tissue.111 Excess FFA will accumulate ectopically, thereby exceeding the rate of fatty acid oxidation and intracellular storage. Ectopic lipid accumulation is the buildup of fat outside normal adipose tissue, such as in the liver and skeletal muscle, which can cause severe IR.111 Ectopic fat produces toxic lipids, for example, ceramide and DAG.112
Lipid-induced IR may occur due to the aggregation of specific lipid mediators. Lipid mediators are lipid molecules that function as intermediaries in cellular signals.113 DAG and ceramide have been widely studied as mediators of lipid-induced IR in the liver and skeletal muscle, able to interfere with insulin signaling pathways.114,115 DAG can accumulate in IR individuals due to an imbalance between its production and the addition of triacylglycerol (TAG) or triglyceride hydrolysis pathways. Ceramide coordinates cellular responses to cytokines or nutritional signals, such as saturated fatty acids.113
DAG is a signal intermediate that activates members of the PKC family. There are three groups in the PKC family: conventional (α, βI, β II, γ), novel (δ, ε, η, θ), and atypical (ζ and λ), and novel PKCs (nPKCs) have a greater affinity for DAG is known to mediate the role of DAG in IR.116 PKC-ε is mainly involved in tissues such as liver and adipose tissue.117 PKC-δ may also be increased in the liver of obese individuals, causing IR through a significant decrease in insulin-stimulated IRS-1 phosphorylation at Tyr612 and a decrease in Akt phosphorylation at Ser473.118 DAG activates PKC-θ in skeletal muscle, leading to IR through increased IRS-1 serine 1101 phosphorylation and inhibition of insulin-stimulated IRS-1 tyrosine phosphorylation and Akt2 phosphorylation.119 Through PKC activation, sn-1,2-DAG mediates lipid-induced insulin resistance in the liver, skeletal muscle, and white adipose tissue (WAT).120,121
Figure 2 illustrates the mechanism of IR in skeletal muscle. Cells take up fatty acids (FA) via a specific transport protein in the cell membrane (CD36).122 Esterification of fatty acids with coenzyme A forms Long Chain Fatty Acid-CoA (LCFA-CoA). When there are too many FAs and/or mitochondrial problems that prevent β-oxidation from occurring, LCFA-CoA builds up and is broken down into DAG and ceramide. DAG induces sustained activation of serine/threonine kinases such as PKC-θ, which facilitates serine phosphorylation of IRS-1. The serine-phosphorylated form of IRS-1 is unable to associate with and activate PI3K, resulting in decreased PI3K and Akt2 activity. GLUT4 regulates glucose transport through cell membranes, which in turn reduces glucose intake and glycogen synthesis due to this decrease in Akt2 activity. The liver’s IR mechanism bears similarities to that of skeletal muscle. When there is more DAG in liver cells, PKC- ε gets activated. This lowers the activity of insulin receptor kinase, which in turn lowers the tyrosine phosphorylation of IRS-2 by insulin. This, in turn, lowers the activation of PI3K and Akt2. When Akt2 activation is minimal, there is a decrease in hepatic glucose uptake. Furthermore, reduced Akt2 activity leads to decreased phosphorylation of the transcription factor forkhead box O (FOXO), allowing it to enter the nucleus and activate transcription of gluconeogenesis rate-controlling enzymes. The result is an increase in liver glucose production. This allows glucose to escape via GLUT2, contributing to increased plasma glucose levels.111,113,123,124
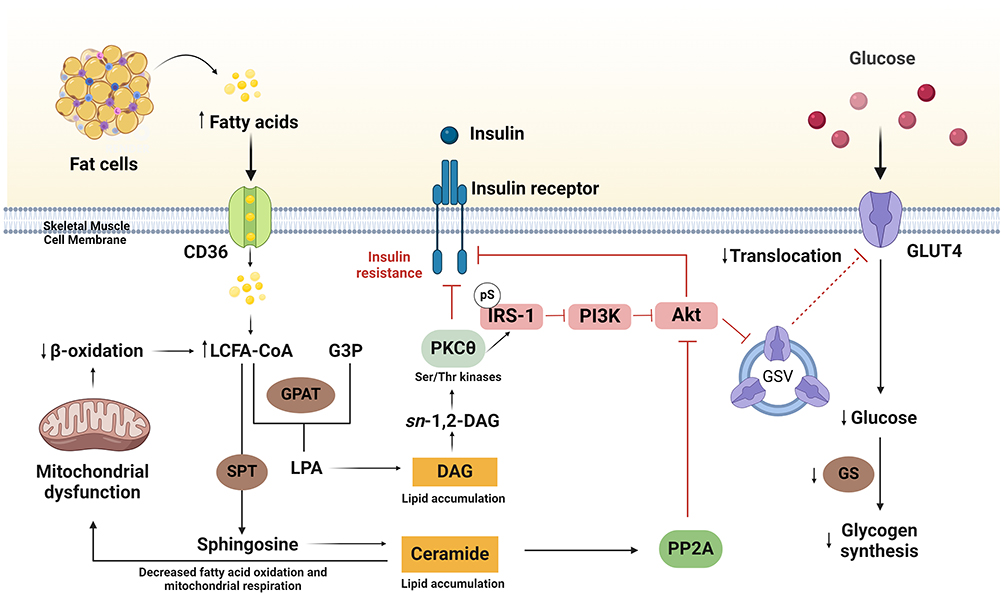 |
Figure 2 The mechanism of insulin resistance (IR) in skeletal muscle. Insulin induces activation of the insulin receptor, resulting in autophosphorylation and activation of the insulin receptor intrinsic tyrosine kinases and subsequently associates with phosphoinositide 3-kinase (PI3K). Activated PI3K will induce protein kinase B (Akt) which contributes a main role in glucose metabolism by regulating the translocation of the cytosolic glucose transporter type 4 (GLUT4), to the skeletal muscle membrane cell. Inhibition of PI3K and/or PKB/Akt attenuates the translocation of GLUT4. Insulin also increases LCFA uptake via a cluster of differentiation 36 (CD36), which leads to the increase of DAG and/or ceramide levels, thus activating PKC and subsequently the occurrence of IR. Created with BioRender.com. Abbreviations: LCFA-CoA, long-chain fatty acid-CoA; DAG, diacylglycerol; PKC, protein kinase C; IR, insulin receptor; IRS, insulin receptor substrate; PI3K, phosphatidylinositol 3-kinase; GLUT4, glucose transport protein 4; G3P, glycerol 3-phosphate; GPAT, glycerol-3-phosphate acyltransferase; LPA, lysophosphatidic acid; SPT, serine palmitoyl transferase; PP2A, protein phosphatase 2A. |
The role of ceramide in inducing IR is still controversial. However, the accumulation of hepatic ceramides and certain dihydroceramides (eg, C16:0) is increased in insulin-resistant humans with non-alcoholic steatohepatitis (NASH) and is positively correlated with oxidative stress and hepatic inflammation.125 Ceramide is a bioactive sphingolipid produced from intracellular fatty acids and sphingosine.111,113 Ceramide disrupts insulin action by activating PP2A, which dephosphorylates Akt2, thereby reducing insulin signaling, but not due to issues in upstream signaling steps like IR or PI3K activation.113 Research shows that liver-specific deletion of Cers6 can reduce ceramide levels in the liver, protect against high-fat diet-induced obesity, and improve glucose tolerance.126
In summary, lipid accumulation and the resulting metabolic disturbances in adipose tissue, liver, and skeletal muscle play an important role in inducing IR through multiple pathways, including activation of inflammatory responses, impaired insulin signaling, and alteration of mitochondrial function.
The Role of Inflammation in Insulin Resistance
Inflammation is a significant factor in the progression of IR in both humans and animals. Elevated levels of proinflammatory cytokines, such as TNF-α and IL-6, are commonly observed in obese people with IR, suggesting the role of inflammatory mechanisms in this illness.127–130 A HFD, in particular, intricately interconnects inflammation and IR. Adipose tissue inflammation in mice that developed obesity due to an HFD begins shortly after the start of the HFD and continues for the duration of the HFD. Nevertheless, the inflammation of adipose tissue decreased promptly following the transition from HFD to a regular diet, suggesting that food significantly influences the regulation of adipose tissue inflammation.131 The mice in the HFD group had a lot more proinflammatory cytokines in their blood when they were at rest. These included TNF-α, IL-6, leptin, MCP-1, plasminogen activator inhibitor-1 (PAI-1), and resistin.132 Research conducted on animal models demonstrates that high-fat diets not only result in elevated levels of proinflammatory cytokines in adipose tissue but also trigger inflammation throughout the entire body. This process entails the activation of the NF-kappaB pathway, which ultimately leads to a decrease in insulin sensitivity. Chiang et al found that giving mice an HFD increased the activation of NF-kappaB. This, in turn, caused higher levels of IKKɛ in both hepatocytes and adipocytes. Furthermore, when IKKɛ was removed in mice, they were shielded from obesity and chronic inflammation caused by the HFD.133
Hotamisligil’s research revealed that pro-inflammatory cytokines can cause IR.134 When there is excess fat, adipose tissue is infiltrated by macrophages and other immune cells, which then produce pro-inflammatory cytokines such as TNF-α, leptin, IL-6, resistin, MCP-1, PAI-1, angiotensinogen, visfatin, retinol binding protein-4, and serum amyloid A. Adipose tissue also produces other substances, such as adiponectin, which decreases as body fat increases.135 Consuming high-fat foods and obesity causes fat to accumulate in fat cells (adipocytes). This buildup triggers stress in cells and activates inflammatory signaling pathways, namely IKKβ/NF-kappaB and JNK. Proinflammatory cytokines like TNF-α and IL-1β turn on the JNK and IKKβ/NF-kappaB pathways through TNFR and IL-R receptor-mediated mechanisms. Cellular stress, such as oxidative stress and endoplasmic reticulum (ER) stress, also activates JNK and NF-kappaB. Lipid accumulation in adipocytes can increase ROS production, which in turn increases TNF-α, IL-6, and MCP-1 production and reduces adiponectin production. Saturated fats trigger an increase in ceramide synthesis, leading to its accumulation in tissues like muscle and a correlation with IR. Acute lipid infusion activates various PKC isoforms that can also activate IKKβ and NF-kappaB.135–137
JNK and IKKβ/NF-kappaB are involved in inflammation-induced IR through different mechanisms. JNK makes IR worse by phosphorylating serine residues in IRS-1. This stops the insulin receptor/IRS-1 pathway from working normally, affecting serine-302 (pS302) and serine-307 (pS307). On the other hand, IKKβ targets IκBα for proteasomal degradation, which allows NF-κB to enter the nucleus and stimulate gene expression. In contrast to JNK, IKKβ induces IR by activating NF-kappaB via transcription, thereby increasing target gene expression and thereby increasing IR.135 Treatments that inhibit NF-kappaB can increase IR, indicating that the NF-κB pathway plays an important role in inflammation-associated IR.138 Furthermore, studies have demonstrated that inhibiting JNK decreases the release of proinflammatory cytokines associated with IR, including TNF- and MCP-1.139,140
The JAK-STAT pathway, especially through IL-6, plays a part in IR by decreasing the expression of GLUT4 and IRS-1 and increasing the expression of SOCS3.141,142 This also stops glycogen synthesis,143 leads to the degradation of IRS-1 by proteasomes, and stops insulin signal transduction.137 IL-6 plays an important role in regulating metabolism and immunity. Humans have shown visceral fat to be an important site for IL-6 secretion.144 A very low-calorie diet and weight loss significantly reduced IL-6 levels in adipose tissue and serum, as well as improved insulin sensitivity compared to controls.145 Additionally, treatment with anti-IL-6 antibodies increased insulin sensitivity in diet-induced obese mice.146
IL-1β is a pro-inflammatory mediator that contributes to IR through multiple mechanisms. It binds to the IL-1R1 receptor, activating the JAK protein kinase that stimulates NF-κB translocation to the nucleus. This triggers the expression of inflammatory genes. IL-1β also reduces IRS-1 tyrosine phosphorylation and disrupts PI3K and Akt signaling.147,148 Experimental studies in animal models indicate that IL-1β is a proinflammatory mediator that can activate various other proinflammatory cytokines and chemokines.149
Obesity-induced inflammation and lipid metabolites contribute significantly to IR through multiple inflammatory and metabolic signaling pathways. Because of this, reducing systemic inflammation and targeting inflammatory signaling pathways and proinflammatory cytokines like TNF-α, IL-6, and IL-1β, may be a good way to fight IR and stop T2DM from happening.
Treatment and Prevention Strategies
It is important to identify gaps in current research regarding the role of lipids in IR. Many studies have linked increased levels of lipids such as DAG and ceramide to IR, but the specific molecular mechanisms of impaired insulin signaling by these lipids remain incompletely understood. For example, Holland et al showed that ceramide synthase inhibitors can increase insulin sensitivity in obese mice.99 Li et al confirmed that a Cordyceps extract with myriocin can lower the buildup of ceramide, boost energy use, and improve glucose homeostasis in obese mice. This suggests that it might be useful as a supplement for treating obesity and related metabolic disorders.150
Dietary intervention for IR is possible. For example, research by Kahleova et al showed that a low-fat plant-based diet significantly reduced body weight, visceral fat, IR, and intramyocellular and hepatocellular lipid levels in overweight adults in 16 weeks.151 Lifestyle interventions and metformin therapy can effectively prevent or delay the disease’s onset. According to research, lifestyle interventions are more effective than metformin, with one case of diabetes prevented per seven people treated over three years.152 It is also important to conduct in-depth investigations into how specific lipids, such as ceramide and DAG, interact with insulin signaling components such as PKC and PP2A, as well as their impact on mitochondria and oxidative stress.
Getting rid of systemic inflammation and targeting inflammatory signaling pathways and proinflammatory cytokines like TNF-α, IL-1β, and IL-6 may help beat IR and stop T2DM from happening. Studies have shown that anti-TNF-α antibodies can improve insulin sensitivity in peripheral tissues and increase adiponectin concentrations.153 Treatment with shRNA can downregulate TNF-α and increase IRS-1 phosphorylation and insulin response.154 IL-1β antibody treatment makes IR, blood sugar control, and β-cell function better in obese mice that were fed a high-fat diet.155,156 Tocilizumab is a humanized monoclonal antibody that selectively blocks IL-6 signaling. This lowers HOMA-IR and IR in RA patients who are not diabetic.157 These studies suggest that inhibiting TNF-α, IL-1β, and IL6 can improve IR and T2DM. In addition, salicylates, an important class of anti-inflammatory agents, have been shown to improve insulin sensitivity and glucose tolerance through inhibition of NF-kappaB and IKKβ.158,159
Omega-3 is a type of polyunsaturated fatty acid that is known to have various health benefits, including positive effects on insulin metabolism and insulin sensitivity. In contrast to saturated fatty acids, which can increase inflammation and contribute to IR, omega-3 reveals anti-inflammatory properties that help reduce the risk of IR. Omega-3 can stimulate the GPR120 receptor, which then inhibits pro-inflammatory signaling pathways in macrophage cells.160 For this process to function, GPR120 must bind to β-arrestin-2. This stops the activation of TAK1, an important kinase in inflammatory pathways.161 By combining findings from human and animal studies, we can gain a more comprehensive understanding of the molecular mechanisms linking inflammation to IR, as well as identify potential interventions to reduce the negative impact of this condition.
Conclusion
IR, a condition associated with various metabolic disorders, is influenced by lipid metabolism, especially induced by HFD. Furthermore, excessive FFA production may lead to decreased insulin sensitivity, progressive pancreatic beta-cell atrophy, and ectopic fat accumulation, which eventually activate inflammatory mediators in the insulin pathway. Human studies show elevated FFAs and TGs may correlate with reduced insulin sensitivity. Metformin and omega-3 fatty acids, or PPAR agonists such as chiglitazar (a novel non-thiazolidinedione structured PPAR pan-agonist) may be potential interventions to overcome these dysfunctions. Animal studies show that HFDs disrupt insulin signaling and increase inflammation, with lipid mediators like ceramides contributing to IR. Understanding lipid-induced IR interactions is critical to developing effective treatments for metabolic diseases. It is suggested to further investigate the long-term impact of dietary changes and pharmacological interventions on lipid metabolism and IR. Future research needs to focus on several specific areas to address these gaps. It will be a challenge to discover new lipid-lowering therapies, specifically those evaluating the efficacy of new compounds that target ceramide and DAG to reduce IR.
Acknowledgments
The authors thank the Directorate of Research and Community Engagement of Padjadjaran University for facilitating the APC.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Wilcox G. Insulin and insulin resistance. Clin Biochem Rev. 2005;26(2):19–39.
2. Al-Goblan AS, Al-Alfi MA, Khan MZ. Mechanism linking diabetes mellitus and obesity. Diabetes Metab Syndr Obes. 2014;7:587–591. doi:10.2147/DMSO.S67400
3. Martins FO, Conde SV. Impact of diet composition on insulin resistance. Nutrients. 2022;14(18):3716. doi:10.3390/nu14183716
4. Perreault L, Newsom SA, Strauss A, et al. Intracellular localization of diacylglycerols and sphingolipids influences insulin sensitivity and mitochondrial function in human skeletal muscle. JCI Insight. 2018;3(3):e96805. doi:10.1172/jci.insight.96805
5. Metcalfe LK, Smith GC, Turner N. Defining lipid mediators of insulin resistance - controversies and challenges. J Mol Endocrinol. 2018. doi:10.1530/JME-18-0023
6. Sokolowska E, Blachnio-Zabielska A. The role of ceramides in Insulin resistance. Front Endocrinol. 2019;10:577. doi:10.3389/fendo.2019.00577
7. Jia W, Ma J, Miao H, et al. Chiglitazar monotherapy with sitagliptin as an active comparator in patients with type 2 diabetes: a randomized, double-blind, phase 3 trial (CMAS). Sci Bull. 2021;66(15):1581–1590. doi:10.1016/j.scib.2021.02.027
8. Ji L, Song W, Fang H, et al. Efficacy and safety of chiglitazar, a novel peroxisome proliferator-activated receptor pan-agonist, in patients with type 2 diabetes: a randomized, double-blind, placebo-controlled, phase 3 trial (CMAP). Sci Bull. 2021;66(15):1571–1580. doi:10.1016/j.scib.2021.03.019
9. Dresner A, Laurent D, Marcucci M, et al. Effects of free fatty acids on glucose transport and IRS-1-associated phosphatidylinositol 3-kinase activity. J Clin Invest. 1999;103(2):253–259. doi:10.1172/JCI5001
10. Paniagua JA. Nutrition, insulin resistance and dysfunctional adipose tissue determine the different components of metabolic syndrome. World J Diabetes. 2016;7(19):483–514. doi:10.4239/wjd.v7.i19.483
11. Shu L, Hou X, Song G, Wang C, Ma H. Comparative analysis of long non-coding RNA expression profiles induced by resveratrol and metformin treatment for hepatic insulin resistance. Int J Mol Med. 2021;48(5):206. doi:10.3892/ijmm.2021.5039
12. Deenadayalan A, Subramanian V, Paramasivan V, et al. Stevioside attenuates insulin resistance in skeletal muscle by facilitating IR/IRS-1/Akt/GLUT 4 signaling pathways: an in vivo and in silico approach. Molecules. 2021;26(24):7689. doi:10.3390/molecules26247689
13. Xia QS, Wu F, Wu WB, et al. Berberine reduces hepatic ceramide levels to improve insulin resistance in HFD-fed mice by inhibiting HIF-2α. Biomed Pharmacother. 2022;150:112955. doi:10.1016/j.biopha.2022.112955
14. Yang X, Xu Y, Gao W, et al. Hyperinsulinemia-induced microglial mitochondrial dynamic and metabolic alterations lead to neuroinflammation in vivo and in vitro. Front Neurosci. 2022;16:1036872. doi:10.3389/fnins.2022.1036872
15. Charytoniuk T, Sztolsztener K, Harasim-Symbor E, et al. Cannabidiol - A phytocannabinoid that widely affects sphingolipid metabolism under conditions of brain insulin resistance. Biomed Pharmacother. 2021;142:112057. doi:10.1016/j.biopha.2021.112057
16. Kim SG, Kim DM, Woo JT, et al. Efficacy and safety of lobeglitazone monotherapy in patients with type 2 diabetes mellitus over 24-weeks: a multicenter, randomized, double-blind, parallel-group, placebo controlled trial. PLoS One. 2014;9(4):e92843. doi:10.1371/journal.pone.0092843
17. Schell J, Betts NM, Foster M, Scofield RH, Basu A. Cranberries improve postprandial glucose excursions in type 2 diabetes. Food Funct. 2017;8(9):3083–3090. doi:10.1039/c7fo00900c
18. Li D, Zhang Y, Liu Y, Sun R, Xia M. Purified anthocyanin supplementation reduces dyslipidemia, enhances antioxidant capacity, and prevents insulin resistance in diabetic patients. J Nutr. 2015;145(4):742–748. doi:10.3945/jn.114.205674
19. Ramanjaneya M, Bensila M, Bettahi I, et al. Dynamic changes in circulating endocrine FGF19 subfamily and fetuin-A in response to intralipid and insulin infusions in healthy and PCOS women. Front Endocrinol. 2020;11:568500. doi:10.3389/fendo.2020.568500
20. Jerobin J, Ramanjaneya M, Bettahi I, et al. Regulation of circulating CTRP-2/CTRP-9 and GDF-8/GDF-15 by intralipids and insulin in healthy control and polycystic ovary syndrome women following chronic exercise training. Lipids Health Dis. 2021;20(1):34. doi:10.1186/s12944-021-01463-3
21. Kahleova H, Hlozkova A, Fleeman R, et al. Fat quantity and quality, as part of a low-fat, vegan diet, are associated with changes in body composition, insulin resistance, and insulin secretion. A 16-week randomized controlled trial. Nutrients. 2019;11(3):615. doi:10.3390/nu11030615
22. Savage DB, Watson L, Carr K, et al. Accumulation of saturated intramyocellular lipid is associated with insulin resistance. J Lipid Res. 2019;60(7):1323–1332. doi:10.1194/jlr.M091942
23. Tricò D, Mengozzi A, Baldi S, et al. Lipid-induced glucose intolerance is driven by impaired glucose kinetics and insulin metabolism in healthy individuals. Metabolism. 2022;134:155247. doi:10.1016/j.metabol.2022.155247
24. Abildgaard J, Henstridge DC, Pedersen AT, et al. In vitro palmitate treatment of myotubes from postmenopausal women leads to ceramide accumulation, inflammation and affected insulin signaling. PLoS One. 2014;9(7):e101555. doi:10.1371/journal.pone.0101555
25. Rondanelli M, Riva A, Petrangolini G, et al. Berberine phospholipid is an effective insulin sensitizer and improves metabolic and hormonal disorders in women with polycystic ovary syndrome: a one-group pretest-post-test explanatory study. Nutrients. 2021;13(10):3665. doi:10.3390/nu13103665
26. Chen Z, Luo J, Jia M, Chai Y, Bao Y. Polygonatum sibiricum saponin exerts beneficial hypoglycemic effects in type 2 diabetes mice by improving hepatic insulin resistance and glycogen synthesis-related proteins. Nutrients. 2022;14(24):5222. doi:10.3390/nu14245222
27. Fan L, Niu H, Zhao L, et al. Purendan alleviates non-alcoholic fatty liver disease in aged type 2 diabetic rats via regulating mTOR/S6K1/SREBP-1c signaling pathway. Biomed Pharmacother. 2022;148:112697. doi:10.1016/j.biopha.2022.112697
28. Zhang Q, Huang Y, Li X, et al. Tangduqing granules attenuate insulin resistance and abnormal lipid metabolism through the coordinated regulation of PPARγ and DGAT2 in type 2 diabetic rats. J Diabetes Res. 2019;2019:7403978. doi:10.1155/2019/7403978
29. Kurtz CL, Peck BC, Fannin EE, et al. MicroRNA-29 fine-tunes the expression of key FOXA2-activated lipid metabolism genes and is dysregulated in animal models of insulin resistance and diabetes. Diabetes. 2014;63(9):3141–3148. doi:10.2337/db13-1015
30. Li X, Jin SJ, Su J, Li XX, Xu M. Acid sphingomyelinase down-regulation alleviates vascular endothelial insulin resistance in diabetic rats. Basic Clin Pharmacol Toxicol. 2018;123(6):645–659. doi:10.1111/bcpt.13073
31. Murtaza M, Khan G, Aftab MF, et al. Cucurbitacin E reduces obesity and related metabolic dysfunction in mice by targeting JAK-STAT5 signaling pathway. PLoS One. 2017;12(6):e0178910. doi:10.1371/journal.pone.0178910
32. Juszczak F, Pierre L, Decarnoncle M, et al. Sex differences in obesity-induced renal lipid accumulation revealed by lipidomics: a role of adiponectin/AMPK axis. Biol Sex Differ. 2023;14(1):63. doi:10.1186/s13293-023-00543-6
33. Manickam R, Tur J, Badole SL, et al. Nampt activator P7C3 ameliorates diabetes and improves skeletal muscle function modulating cell metabolism and lipid mediators. J Cachexia, Sarcopenia Muscle. 2022;13(2):1177–1196. doi:10.1002/jcsm.12887
34. Kim Y, Lim JH, Kim EN, et al. Adiponectin receptor agonist ameliorates cardiac lipotoxicity via enhancing ceramide metabolism in type 2 diabetic mice. Cell Death Dis. 2022;13(3):282. doi:10.1038/s41419-022-04726-8
35. Hosseini Dastgerdi A, Sharifi M, Soltani N. GABA administration improves liver function and insulin resistance in offspring of type 2 diabetic rats. Sci Rep. 2021;11(1):23155. doi:10.1038/s41598-021-02324-w
36. Elzinga SE, Savelieff MG, O’Brien PD, et al. Sex differences in insulin resistance, but not peripheral neuropathy, in a diet-induced prediabetes mouse model. Dis Model Mech. 2021;14(4):dmm048909. doi:10.1242/dmm.048909
37. Agostino M, Rooney J, Herat L, et al. TNFSF14-Derived molecules as a novel treatment for obesity and type 2 diabetes. Int J Mol Sci. 2021;22(19):10647. doi:10.3390/ijms221910647
38. Chen Y, Hu W, Li Q, et al. NGBR is required to ameliorate type 2 diabetes in mice by enhancing insulin sensitivity. J Biol Chem. 2021;296:100624. doi:10.1016/j.jbc.2021.100624
39. Obayemi MJ, Akintayo CO, Oniyide AA, et al. Protective role of melatonin against adipose-hepatic metabolic comorbidities in experimentally induced obese rat model. PLoS One. 2021;16(12):e0260546. doi:10.1371/journal.pone.0260546
40. Ali ES, Girard D, Petrovsky N. Impaired Ca2+ signaling due to hepatic steatosis mediates hepatic insulin resistance in Alström syndrome mice that is reversed by GLP-1 analog treatment. Am J Physiol Cell Physiol. 2021;321(1):C187–C198. doi:10.1152/ajpcell.00020.2021
41. Zeng Q, Wang N, Zhang Y, et al. Partial deficiency of Zfp217 resists high-fat diet-induced obesity by increasing energy metabolism in mice. Int J Mol Sci. 2021;22(10):5390. doi:10.3390/ijms22105390
42. Liu Y, Qiu Y, Chen Q, Han X, Cai M, Hao L. Puerarin suppresses the hepatic gluconeogenesis via activation of PI3K/Akt signaling pathway in diabetic rats and HepG2 cells. Biomed Pharmacother. 2021;137:111325. doi:10.1016/j.biopha.2021.111325
43. Liu X, Chen S, Zhang L. Downregulated microRNA-130b-5p prevents lipid accumulation and insulin resistance in a murine model of nonalcoholic fatty liver disease. Am J Physiol Endocrinol Metab. 2020;319(1):E34–E42. doi:10.1152/ajpendo.00528.2019
44. Fang K, Gu M. Crocin Improves insulin sensitivity and ameliorates adiposity by regulating AMPK-CDK5-PPARγ signaling. Biomed Res Int. 2020;2020:9136282. doi:10.1155/2020/9136282
45. Yagi T, Ataka K, Cheng KC, et al. Red rice koji extract alleviates hyperglycemia by increasing glucose uptake and glucose transporter type 4 levels in skeletal muscle in two diabetic mouse models. Food Nutr Res. 2020;64. doi:10.29219/fnr.v64.4226
46. Wohua Z, Weiming X. Glutaredoxin 2 (GRX2) deficiency exacerbates high fat diet (HFD)-induced insulin resistance, inflammation and mitochondrial dysfunction in brain injury: a mechanism involving GSK-3β. Biomed Pharmacother. 2019;118:108940. doi:10.1016/j.biopha.2019.108940
47. Purvis GSD, Collino M, Loiola RA, et al. Identification of annexinA1 as an endogenous regulator of RhoA, and its role in the pathophysiology and experimental therapy of type-2 diabetes. Front Immunol. 2019;10:571. doi:10.3389/fimmu.2019.00571
48. Ohkura T, Yoshimura T, Fujisawa M, et al. Spred2 regulates high fat diet-induced adipose tissue inflammation, and metabolic abnormalities in mice. Front Immunol. 2019;10:17. doi:10.3389/fimmu.2019.00017
49. Gao H, Guo Y, Yan Q, et al. Lipoatrophy and metabolic disturbance in mice with adipose-specific deletion of kindlin-2. JCI Insight. 2019;4(13):e128405. doi:10.1172/jci.insight.128405
50. Aarts S, Reiche M, den Toom M, et al. Depletion of CD40 on CD11c+ cells worsens the metabolic syndrome and ameliorates hepatic inflammation during NASH. Sci Rep. 2019;9(1):14702. doi:10.1038/s41598-019-50976-6
51. Bai YF, Wang SW, Wang XX, et al. The flavonoid-rich Quzhou Fructus Aurantii extract modulates gut microbiota and prevents obesity in high-fat diet-fed mice. Nutr Diabetes. 2019;9(1):30. doi:10.1038/s41387-019-0097-6
52. Ren X, Chen N, Chen Y, Liu W, Hu Y. TRB3 stimulates SIRT1 degradation and induces insulin resistance by lipotoxicity via COP1. Exp Cell Res. 2019;382(1):111428. doi:10.1016/j.yexcr.2019.05.009
53. Murano K, Ogino H, Okuno T, Arakawa T, Ueno H. Role of supplementary selenium on the induction of insulin resistance and oxidative stress in NSY mice fed a high fat diet. Biol Pharm Bull. 2018;41(1):92–98. doi:10.1248/bpb.b17-00622
54. Rico JE, Giesy SL, Haughey NJ, et al. Intravenous triacylglycerol infusion promotes ceramide accumulation and hepatic steatosis in dairy cows. J Nutr. 2018;148(10):1529–1535. doi:10.1093/jn/nxy155
55. Meng F, Lin Y, Yang M, et al. JAZF1 inhibits adipose tissue macrophages and adipose tissue inflammation in diet-Induced diabetic mice. Biomed Res Int. 2018;2018:4507659. doi:10.1155/2018/4507659
56. Hossain Z, Valicherla GR, Gupta AP, et al. Discovery of pancreastatin inhibitor PSTi8 for the treatment of insulin resistance and diabetes: studies in rodent models of diabetes mellitus. Sci Rep. 2018;8(1):8715. doi:10.1038/s41598-018-27018-8
57. Alwahsh SM, Dwyer BJ, Forbes S, et al. Insulin production and resistance in different models of diet-induced obesity and metabolic syndrome. Int J Mol Sci. 2017;18(2):285. doi:10.3390/ijms18020285
58. Guo H, Wang B, Li H, Ling L, Niu J, Gu Y. Glucagon-like peptide-1 analog prevents obesity-related glomerulopathy by inhibiting excessive autophagy in podocytes. Am J Physiol Renal Physiol. 2018;314(2):F181–F189. PMID: 29070572. doi:10.1152/ajprenal.00302.2017
59. Niu M, Xiang L, Liu Y, et al. Adiponectin induced AMP-activated protein kinase impairment mediates insulin resistance in Bama mini-pig fed high-fat and high-sucrose diet. Asian-Australas J Anim Sci. 2017;30(8):1190–1197. doi:10.5713/ajas.17.0006
60. Mannerås-Holm L, Schönke M, Brozinick JT, et al. Diacylglycerol kinase ε deficiency preserves glucose tolerance and modulates lipid metabolism in obese mice. J Lipid Res. 2017;58(5):907–915. doi:10.1194/jlr.M074443
61. Zhang L, Du J, Yano N, et al. Sodium butyrate protects -against high fat diet-induced cardiac dysfunction and metabolic disorders in type II diabetic mice. J Cell Biochem. 2017;118(8):2395–2408. doi:10.1002/jcb.25902
62. Zhang W, Wu M, Kim T, et al. Skeletal muscle TRIB3 mediates glucose toxicity in diabetes and high-fat diet-induced insulin resistance. Diabetes. 2016;65(8):2380–2391. doi:10.2337/db16-0154
63. Li GS, Liu XH, Zhu H, et al. Skeletal muscle insulin resistance in hamsters with diabetes developed from obesity is involved in abnormal skeletal muscle LXR, PPAR and SREBP expression. Exp Ther Med. 2016;11(6):2259–2269. doi:10.3892/etm.2016.3209
64. Kieswich J, Sayers SR, Silvestre MF, et al. Monomeric eNAMPT in the development of experimental diabetes in mice: a potential target for type 2 diabetes treatment. Diabetologia. 2016;59(11):2477–2486. doi:10.1007/s00125-016-4076-3
65. Shi CX, Zhao MX, Shu XD, et al. β-aminoisobutyric acid attenuates hepatic endoplasmic reticulum stress and glucose/lipid metabolic disturbance in mice with type 2 diabetes. Sci Rep. 2016;6:21924. doi:10.1038/srep21924
66. van Diepen JA, Jansen PA, Ballak DB, et al. Genetic and pharmacological inhibition of vanin-1 activity in animal models of type 2 diabetes. Sci Rep. 2016;6:21906. doi:10.1038/srep21906
67. Tao W, Wu J, Xie BX, et al. Lipid-induced muscle insulin resistance is mediated by GGPPS via modulation of the RhoA/Rho kinase signaling pathway. J Biol Chem. 2015;290(33):20086–20097. doi:10.1074/jbc.M115.657742
68. Lu Z, Zhang X, Li Y, et al. TLR4 antagonist attenuates atherogenesis in LDL receptor-deficient mice with diet-induced type 2 diabetes. Immunobiology. 2015;220(11):1246–1254. doi:10.1016/j.imbio.2015.06.016
69. Li Y, Lu Z, Zhang X, et al. Metabolic syndrome exacerbates inflammation and bone loss in periodontitis. J Dent Res. 2015;94(2):362–370. doi:10.1177/0022034514561658
70. Stenkula KG, Lindahl M, Petrlova J, et al. Single injections of apoA-I acutely improve in vivo glucose tolerance in insulin-resistant mice. Diabetologia. 2014;57(4):797–800. doi:10.1007/s00125-014-3162-7
71. Siersbæk MS, Ditzel N, Hejbøl EK, et al. C57BL/6J substrain differences in response to high-fat diet intervention. Sci Rep. 2020;10(1):14052. doi:10.1038/s41598-020-70765-w
72. Magliano D, Boyko EJ. IDF Diabetes Atlas.
73. World Health Organization. Diabetes; 2023. Available from: https://www.who.int/health-topics/diabetes.
74. Mukhtar Y, Galalain A, Yunusa U. A modern overview on diabetes mellitus: a chronic endocrine disorder. Europ J Biol. 2020;5(2):1–14.518. doi:10.47672/ejb.409
75. Galicia-Garcia U, Benito-Vicente A, Jebari S, et al. Pathophysiology of type 2 diabetes mellitus. Int J Mol Sci. 2020;21(17):6275. doi:10.3390/ijms21176275
76. Yaribeygi H, Farrokhi FR, Butler AE, Sahebkar A. Insulin resistance: review of the underlying molecular mechanisms. J Cell Physiol. 2019;234(6):8152–8161. doi:10.1002/jcp.27603
77. Ahmed B, Sultana R, Greene MW. Adipose tissue and insulin resistance in obese. Biomed Pharmacother. 2021;137:111315. doi:10.1016/j.biopha.2021.111315
78. Lehrke M, Marx N. Diabetes mellitus and heart failure. Am J Med. 2017;130(6S):S40–S50. doi:10.1016/j.amjmed.2017.04.010
79. Lau LH, Lew J, Borschmann K, Thijs V, Ekinci EI. Prevalence of diabetes and its effects on stroke outcomes: a meta-analysis and literature review. J Diabetes Investig. 2019;10(3):780–792. doi:10.1111/jdi.12932
80. Jia G, Sowers JR. Hypertension in diabetes: an update of basic mechanisms and clinical disease. Hypertension. 2021;78(5):1197–1205. doi:10.1161/HYPERTENSIONAHA.121.17981
81. Kojta I, Chacińska M, Błachnio-Zabielska A. Obesity, bioactive lipids, and adipose tissue inflammation in insulin resistance. Nutrients. 2020;12(5):1305. doi:10.3390/nu12051305
82. Wondmkun YT. Obesity, insulin resistance, and type 2 diabetes: associations and therapeutic Implications. Diabetes Metab Syndr Obes. 2020;13:3611–3616. doi:10.2147/DMSO.S275898
83. Lin X, Li H. Obesity: epidemiology, Pathophysiology, and Therapeutics. Front Endocrinol. 2021;12:706978. doi:10.3389/fendo.2021.706978
84. Al-Jawaldeh A, Abbass MMS. Unhealthy dietary habits and obesity: the major risk factors beyond non-communicable diseases in the eastern Mediterranean region. Front Nutr. 2022;9:817808. doi:10.3389/fnut.2022.817808
85. White U, Beyl RA, Ravussin E. A higher proportion of small adipocytes is associated with increased visceral and ectopic lipid accumulation during weight gain in response to overfeeding in men. Int J Obes Lond. 2022;46(8):1560–1563. doi:10.1038/s41366-022-01150-y
86. Longo M, Zatterale F, Naderi J, et al. Adipose tissue dysfunction as determinant of obesity-associated metabolic complications. Int J Mol Sci. 2019;20(9):2358. doi:10.3390/ijms20092358
87. Ghosh A, Gao L, Thakur A, Siu PM, Lai CWK. Role of free fatty acids in endothelial dysfunction. J Biomed Sci. 2017;24(1):50. doi:10.1186/s12929-017-0357-5
88. Galaviz KI, Narayan KMV, Lobelo F, Weber MB. Lifestyle and the prevention of type 2 diabetes: a status report. Am J Lifestyle Med. 2015;12(1):4–20. doi:10.1177/1559827615619159
89. Asif M. The prevention and control the type-2 diabetes by changing lifestyle and dietary pattern. J Educ Health Promot. 2014;3:1. doi:10.4103/2277-9531.127541
90. Gonzalez-Lopez C, Wojeck BS. Role of metformin in the management of type 2 diabetes: recent advances. Pol Arch Intern Med. 2023;133(6):16511. doi:10.20452/pamw.16511
91. Chacińska M, Zabielski P, Książek M, et al. The impact of omega-3 fatty acids supplementation on insulin resistance and content of adipocytokines and biologically active lipids in adipose tissue of high-fat diet fed rats. Nutrients. 2019;11(4):835. doi:10.3390/nu11040835
92. Kalupahana NS, Claycombe KJ, Moustaid-Moussa N. (n-3) Fatty acids alleviate adipose tissue inflammation and insulin resistance: mechanistic insights. Adv Nutr. 2011;2(4):304–316. doi:10.3945/an.111.000505
93. Kahn D, Perreault L, Macias E, et al. Subcellular localisation and composition of intramuscular triacylglycerol influence insulin sensitivity in humans. Diabetologia. 2021;64(1):168–180. doi:10.1007/s00125-020-05315-0
94. Pickersgill L, Litherland GJ, Greenberg AS, Walker M, Yeaman SJ. Key role for ceramides in mediating insulin resistance in human muscle cells. J Biol Chem. 2007;282(17):12583–12589. doi:10.1074/jbc.M611157200
95. Manco M, Mingrone G, Greco AV, et al. Insulin resistance directly correlates with increased saturated fatty acids in skeletal muscle triglycerides. Metabolism. 2000;49(2):220–224. doi:10.1016/s0026-0495(00)91377-5
96. Muoio DM. Intramuscular triacylglycerol and insulin resistance: guilty as charged or wrongly accused? Biochim Biophys Acta. 2010;1801(3):281–288. doi:10.1016/j.bbalip.2009.11.007
97. Adams JM, Pratipanawatr T, Berria R, et al. Ceramide content is increased in skeletal muscle from obese insulin-resistant humans. Diabetes. 2004;53(1):25–31. doi:10.2337/diabetes.53.1.25
98. Summers SA. Ceramides in insulin resistance and lipotoxicity. Prog Lipid Res. 2006;45(1):42–72. doi:10.1016/j.plipres.2005.11.002
99. Holland WL, Brozinick JT, Wang LP, et al. Inhibition of ceramide synthesis ameliorates glucocorticoid-, saturated-fat-, and obesity-induced insulin resistance. Cell Metab. 2007;5(3):167–179. doi:10.1016/j.cmet.2007.01.002
100. Ofori EK, Buabeng A, Amanquah SD, et al. Effect of circulating ceramides on adiposity and insulin resistance in patients with type 2 diabetes: an observational cross-sectional study. Endocrinol Diabetes Metab. 2023;6(3):e418. doi:10.1002/edm2.418
101. Haus JM, Kashyap SR, Kasumov T, et al. Plasma ceramides are elevated in obese subjects with type 2 diabetes and correlate with the severity of insulin resistance. Diabetes. 2009;58(2):337–343. doi:10.2337/db08-1228
102. Broskey NT, Obanda DN, Burton JH, Cefalu WT, Ravussin E. Skeletal muscle ceramides and daily fat oxidation in obesity and diabetes. Metabolism. 2018;82:118–123. doi:10.1016/j.metabol.2017.12.012
103. Chiazza F, Couturier-Maillard A, Benetti E, et al. Targeting the NLRP3 inflammasome to reduce diet-induced metabolic abnormalities in mice. Mol Med. 2016;21(1):1025–1037. doi:10.2119/molmed.2015.00104
104. Huang Y, Zhou T, Zhang Y, et al. Antidiabetic activity of a Flavonoid-Rich extract from flowers of Wisteria sinensis in type 2 diabetic mice via activation of the IRS-1/PI3K/Akt/GLUT4 pathway. J Funct Foods. 2021;77:104338. doi:10.1016/j.jff.2020.104338
105. Van Beek M, Oravecz-Wilson KI, Delekta PC, et al. Bcl10 links saturated fat overnutrition with hepatocellular NF-kB activation and insulin resistance. Cell Rep. 2012;1(5):444–452. doi:10.1016/j.celrep.2012.04.006
106. Braunersreuther V, Viviani GL, Mach F, Montecucco F. Role of cytokines and chemokines in non-alcoholic fatty liver disease. World J Gastroenterol. 2012;18(8):727–735. PMID: 22371632; PMCID: PMC3286135. doi:10.3748/wjg.v18.i8.727
107. Kanda H, Tateya S, Tamori Y, et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J Clin Invest. 2006;116(6):1494–1505. doi:10.1172/JCI26498
108. Manandhar B, Ahn JM. Glucagon-like peptide-1 (GLP-1) analogs: recent advances, new possibilities, and therapeutic implications. J Med Chem. 2015;58(3):1020–1037. doi:10.1021/jm500810s
109. Müller TD, Finan B, Bloom SR, et al. Glucagon-like peptide 1 (GLP-1). Mol Metab. 2019;30:72–130. doi:10.1016/j.molmet.2019.09.010
110. Nauck MA, Quast DR, Wefers J, Meier JJ. GLP-1 receptor agonists in the treatment of type 2 diabetes - state-of-the-art. Mol Metab. 2021;46:101102. doi:10.1016/j.molmet.2020.101102
111. Chandrasekaran P, Weiskirchen R. Cellular and molecular mechanisms of insulin resistance. Curr Tissue Microenviron Rep. 2024. doi:10.1007/s43152-024-00056-3
112. Vazquez-Jimenez JG, Roura A, Jimenez-Mena L, Olivares-Reyes J. Role of free fatty acids on insulin resistance. Gaceta medica de Mexico. 2018;153. doi:10.24875/GMM.M18000092
113. Samuel VT, Shulman GI. Mechanisms for insulin resistance: common threads and missing links. Cell. 2012;148(5):852–871. doi:10.1016/j.cell.2012.02.017
114. Mastrototaro L, Roden M. Insulin resistance and insulin sensitizing agents. Metabolism. 2021;125:154892. doi:10.1016/j.metabol.2021.154892
115. Gancheva S, Jelenik T, Álvarez-Hernández E, Roden M. Interorgan Metabolic crosstalk in human insulin resistance. Physiol Rev. 2018;98(3):1371–1415. doi:10.1152/physrev.00015.2017
116. Lee SH, Park SY, Choi CS. Insulin resistance: from mechanisms to therapeutic strategies. Diabetes Metab J. 2022;46(1):15–37. doi:10.4093/dmj.2021.0280
117. Lyu K, Zhang D, Song J, et al. Short-term overnutrition induces white adipose tissue insulin resistance through sn-1,2-diacylglycerol/PKCε/insulin receptor Thr1160 phosphorylation. JCI Insight. 2021;6(4):e139946. doi:10.1172/jci.insight.139946
118. Bezy O, Tran TT, Pihlajamäki J, et al. PKCδ regulates hepatic insulin sensitivity and hepatosteatosis in mice and humans. J Clin Invest. 2011;121(6):2504–2517. doi:10.1172/JCI46045
119. Szendroedi J, Yoshimura T, Phielix E, et al. Role of diacylglycerol activation of PKCθ in lipid-induced muscle insulin resistance in humans. Proc Natl Acad Sci U S A. 2014;111(26):9597–9602. doi:10.1073/pnas.1409229111
120. Gaspar RC, Lyu K, Hubbard BT, et al. Distinct subcellular localisation of intramyocellular lipids and reduced PKCε/PKCθ activity preserve muscle insulin sensitivity in exercise-trained mice. Diabetologia. 2023;66(3):567–578. doi:10.1007/s00125-022-05838-8
121. Jornayvaz FR, Shulman GI. Diacylglycerol activation of protein kinase Cε and hepatic insulin resistance. Cell Metab. 2012;15(5):574–584. doi:10.1016/j.cmet.2012.03.005
122. Glatz JF, Bonen A, Luiken JJ. Exercise and insulin increase muscle fatty acid uptake by recruiting putative fatty acid transporters to the sarcolemma. Curr Opin Clin Nutr Metab Care. 2002;5(4):365–370. doi:10.1097/00075197-200207000-00003
123. Morino K, Petersen KF, Shulman GI. Molecular mechanisms of insulin resistance in humans and their potential links with mitochondrial dysfunction. Diabetes. 2006;55:S9–S15. PMID: 17130651; PMCID: PMC2995546. doi:10.2337/db06-S002
124. Snel M, Jonker JT, Schoones J, et al. Ectopic fat and insulin resistance: pathophysiology and effect of diet and lifestyle interventions. Int J Endocrinol. 2012;2012:983814. doi:10.1155/2012/983814
125. Apostolopoulou M, Gordillo R, Koliaki C, et al. Specific hepatic sphingolipids relate to insulin resistance, oxidative stress, and inflammation in nonalcoholic steatohepatitis. Diabetes Care. 2018;41(6):1235–1243. doi:10.2337/dc17-1318
126. Turpin SM, Nicholls HT, Willmes DM, et al. Obesity-induced CerS6-dependent C16:0 ceramide production promotes weight gain and glucose intolerance. Cell Metab. 2014;20(4):678–686. doi:10.1016/j.cmet.2014.08.002
127. Almuraikhy S, Kafienah W, Bashah M, et al. Interleukin-6 induces impairment in human subcutaneous adipogenesis in obesity-associated insulin resistance. Diabetologia. 2016;59(11):2406–2416. doi:10.1007/s00125-016-4031-3
128. Mohallem R, Aryal UK. Regulators of TNFα mediated insulin resistance elucidated by quantitative proteomics. Sci Rep. 2020;10(1):20878. doi:10.1038/s41598-020-77914-1
129. Al-Mansoori L, Al-Jaber H, Prince MS, Elrayess MA. Role of inflammatory cytokines, growth factors and adipokines in adipogenesis and insulin resistance. Inflammation. 2022;45(1):31–44. doi:10.1007/s10753-021-01559-z
130. Goyal R, Faizy AF, Siddiqui SS, Singhai M. Evaluation of TNF-α and IL-6 levels in obese and non-obese diabetics: pre- and postinsulin effects. N Am J Med Sci. 2012;4(4):180–184. doi:10.4103/1947-2714.94944
131. Wu H, Ballantyne CM. Metabolic inflammation and insulin resistance in obesity. Circulation Research. 2020;126(11):1549–1564. doi:10.1161/CIRCRESAHA.119.315896
132. Avtanski D, Pavlov VA, Tracey KJ, Poretsky L. Characterization of inflammation and insulin resistance in high-fat diet-induced male C57BL/6J mouse model of obesity. Animal Model Exp Med. 2019;2(4):252–258. doi:10.1002/ame2.12084
133. Chiang SH, Bazuine M, Lumeng CN, et al. The protein kinase IKKepsilon regulates energy balance in obese mice. Cell. 2009;138(5):961–975. doi:10.1016/j.cell.2009.06.046
134. Hotamisligil GS, Shargill NS, Spiegelman BM. Adipose expression of tumor necrosis factor-alpha: direct role in obesity-linked insulin resistance. Science. 1993;259(5091):87–91. doi:10.1126/science.7678183
135. Shoelson SE, Lee J, Goldfine AB. Inflammation and insulin resistance. J Clin Invest. 2006;116(7):1793–1801. doi:10.1172/JCI29069
136. Chen L, Chen R, Wang H, Liang F. Mechanisms linking inflammation to insulin resistance. Int J Endocrinol. 2015;2015:508409. doi:10.1155/2015/508409
137. Li H, Meng Y, He S, et al. Macrophages, chronic inflammation, and insulin resistance. Cells. 2022;11(19):3001. doi:10.3390/cells11193001
138. Yekollu SK, Thomas R, O’Sullivan B. Targeting curcusomes to inflammatory dendritic cells inhibits NF-κB and improves insulin resistance in obese mice. Diabetes. 2011;60(11):2928–2938. PMID: 21885868; PMCID: PMC3198103. doi:10.2337/db11-0275
139. Wu X, Mi Y, Yang H, Hu A, Zhang Q, Shang C. The activation of HMGB1 as a progression factor on inflammation response in normal human bronchial epithelial cells through RAGE/JNK/NF-κB pathway. Mol Cell Biochem. 2013;380(1–2):249–257. doi:10.1007/s11010-013-1680-0
140. Jin SM, Kim KS, Lee SY, et al. The sequential combination of a JNK inhibitor and simvastatin protects porcine islets from peritransplant apoptosis and inflammation. Cell Transplant. 2011;20(7):1139–1151. doi:10.3727/096368910X550170
141. Lukic L, Lalic NM, Rajkovic N, et al. Hypertension in obese type 2 diabetes patients is associated with increases in insulin resistance and IL-6 cytokine levels: potential targets for an efficient preventive intervention. Int J Environ Res Public Health. 2014;11(4):3586–3598. PMID: 24686488; PMCID: PMC4025026. doi:10.3390/ijerph110403586
142. Serrano-Marco L, Barroso E, El kochairi I, et al. The peroxisome proliferator-activated receptor (PPAR) β/δ agonist GW501516 inhibits IL-6-induced signal transducer and activator of transcription 3 (STAT3) activation and insulin resistance in human liver cells. Diabetologia. 2012;55(3):743–751. doi:10.1007/s00125-011-2401-4
143. Dou L, Zhao T, Wang L, et al. miR-200s contribute to interleukin-6 (IL-6)-induced insulin resistance in hepatocytes. J Biol Chem. 2013;288(31):22596–22606. doi:10.1074/jbc.M112.423145
144. Fontana L, Eagon JC, Trujillo ME, Scherer PE, Klein S. Visceral fat adipokine secretion is associated with systemic inflammation in obese humans. Diabetes. 2007;56(4):1010–1013. doi:10.2337/db06-1656
145. Bastard JP, Jardel C, Bruckert E, et al. Elevated levels of interleukin 6 are reduced in serum and subcutaneous adipose tissue of obese women after weight loss. J Clin Endocrinol Metab. 2000;85(9):3338–3342. PMID: 10999830. doi:10.1210/jcem.85.9.6839
146. Klover PJ, Clementi AH, Mooney RA. Interleukin-6 depletion selectively improves hepatic insulin action in obesity. Endocrinology. 2005;146(8):3417–3427. doi:10.1210/en.2004-1468
147. Rehman K, Akash MS. Mechanisms of inflammatory responses and development of insulin resistance: how are they interlinked? J Biomed Sci. 2016;23(1):87. doi:10.1186/s12929-016-0303-y
148. Peiró C, Lorenzo Ó, Carraro R, Sánchez-Ferrer CF. IL-1β inhibition in cardiovascular complications associated to diabetes mellitus. Front Pharmacol. 2017;8:363. doi:10.3389/fphar.2017.00363
149. Akash MS, Shen Q, Rehman K, Chen S. Interleukin-1 receptor antagonist: a new therapy for type 2 diabetes mellitus. J Pharm Sci. 2012;101(5):1647–1658. doi:10.1002/jps.23057
150. Li Y, Talbot CL, Chandravanshi B, et al. Cordyceps inhibits ceramide biosynthesis and improves insulin resistance and hepatic steatosis. Sci Rep. 2022;12(1):7273. doi:10.1038/s41598-022-11219-3
151. Kahleova H, Petersen KF, Shulman GI, et al. Effect of a low-fat vegan diet on body weight, insulin sensitivity, postprandial metabolism, and intramyocellular and hepatocellular lipid levels in overweight adults: a randomized clinical trial. JAMA Network Open. 2020;3(11):e2025454. doi:10.1001/jamanetworkopen.2020.25454
152. Knowler WC, Barrett-Connor E, Fowler SE, et al. Reduction in the incidence of type 2 diabetes with lifestyle intervention or metformin. N Engl J Med. 2002;346(6):393–403. PMID: 11832527; PMCID: PMC1370926. doi:10.1056/NEJMoa012512
153. Lo J, Bernstein LE, Canavan B, et al. Effects of TNF-alpha neutralization on adipocytokines and skeletal muscle adiposity in the metabolic syndrome. Am J Physiol Endocrinol Metab. 2007;293(1):E102–9. doi:10.1152/ajpendo.00089.2007
154. Alipourfard I, Datukishvili N, Mikeladze D. TNF-α downregulation modifies insulin receptor substrate 1 (IRS-1) in metabolic signaling of diabetic insulin-resistant hepatocytes. Mediators Inflamm. 2019;2019:3560819. doi:10.1155/2019/3560819
155. Osborn O, Brownell SE, Sanchez-Alavez M, et al. Treatment with an Interleukin 1 beta antibody improves glycemic control in diet-induced obesity. Cytokine. 2008;44(1):141–148. doi:10.1016/j.cyto.2008.07.004
156. Akash MS, Rehman K, Sun H, Chen S. Sustained delivery of IL-1Ra from PF127-gel reduces hyperglycemia in diabetic GK-rats. PLoS One. 2013;8(2):e55925. doi:10.1371/journal.pone.0055925
157. Schultz O, Oberhauser F, Saech J, et al. Effects of inhibition of interleukin-6 signalling on insulin sensitivity and lipoprotein (a) levels in human subjects with rheumatoid diseases. PLoS One. 2010;5(12):e14328. doi:10.1371/journal.pone.0014328
158. Fleischman A, Shoelson SE, Bernier R, Goldfine AB. Salsalate improves glycemia and inflammatory parameters in obese young adults. Diabetes Care. 2008;31(2):289–294. doi:10.2337/dc07-1338
159. Hundal RS, Petersen KF, Mayerson AB, et al. Mechanism by which high-dose aspirin improves glucose metabolism in type 2 diabetes. J Clin Invest. 2002;109(10):1321–1326. doi:10.1172/JCI14955
160. Glass CK, Olefsky JM. Inflammation and lipid signaling in the etiology of insulin resistance. Cell Metab. 2012;15(5):635–645. doi:10.1016/j.cmet.2012.04.001
161. Oh DY, Talukdar S, Bae EJ, et al. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell. 2010;142(5):687–698. doi:10.1016/j.cell.2010.07.041
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Lingguizhugan Decoction Targets Intestinal Microbiota and Metabolites to Reduce Insulin Resistance in High-Fat Diet Rats

Ning Y, Gong Y, Zheng T, Xie Y, Yuan S, Ding W
Diabetes, Metabolic Syndrome and Obesity 2022, 15:2427-2442
Published Date: 9 August 2022
Tang-Ping-San Decoction Remodel Intestinal Flora and Barrier to Ameliorate Type 2 Diabetes Mellitus in Rodent Model
Yin W, Zhang SQ, Pang WL, Chen XJ, Wen J, Hou J, Wang C, Song LY, Qiu ZM, Liang PT, Yuan JL, Yang ZS, Bian Y
Diabetes, Metabolic Syndrome and Obesity 2022, 15:2563-2581
Published Date: 20 August 2022
Relationship Between Acyl and Desacyl Ghrelin Levels with Insulin Resistance and Body Fat Mass in Type 2 Diabetes Mellitus
Zang P, Yang CH, Liu J, Lei HY, Wang W, Guo QY, Lu B, Shao JQ
Diabetes, Metabolic Syndrome and Obesity 2022, 15:2763-2770
Published Date: 7 September 2022
Association Between Triglyceride-Glucose Index and Serum Uric Acid Levels: A Biochemical Study on Anthropometry in Non-Obese Type 2 Diabetes Mellitus Patients
Luo Y, Hao J, He X, Wang C, Zhao H, Zhang Z, Yang L, Ren L
Diabetes, Metabolic Syndrome and Obesity 2022, 15:3447-3458
Published Date: 3 November 2022
The Association Between Sarcopenia and Diabetes: From Pathophysiology Mechanism to Therapeutic Strategy
Chen H, Huang X, Dong M, Wen S, Zhou L, Yuan X
Diabetes, Metabolic Syndrome and Obesity 2023, 16:1541-1554
Published Date: 30 May 2023