")
Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 18
Investigating the Impact of Circulating Immune Cells on Acne Using Mendelian Randomization
Authors Wang X , Guan S, Xia Z, Zhang X, Bai Y, Li Q, Li H, Yang R
Received 25 March 2025
Accepted for publication 27 June 2025
Published 8 July 2025 Volume 2025:18 Pages 1665—1675
DOI https://doi.org/10.2147/CCID.S525621
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Anne-Claire Fougerousse
Xudong Wang,1,2,* Shaoyu Guan,3,* Zhikuan Xia,2 Xin Zhang,2 Yu Bai,2 Qiang Li,4 Haitao Li,2 Rongya Yang2
1Southern Medical District of Chinese PLA General Hospital, Beijing, 100843, People’s Republic of China; 2Department of Dermatology, Seventh Medical Center of Chinese PLA General Hospital, Beijing, 100010, People’s Republic of China; 3Pharmaceutical Sciences Research Division, Department of Pharmacy, Medical Supplies Centre of PLA General Hospital, Beijing, 100853, China; Medical School of Chinese PLA, Beijing, 100853, People’s Republic of China; 4Medical Health Care Department, Air Force Medical Center PLA, Beijing, 100142, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Rongya Yang, Department of Dermatology, Seventh Medical Center of Chinese PLA General Hospital, Beijing, 100010, People’s Republic of China, Email [email protected] Haitao Li, Department of Dermatology, Seventh Medical Center of Chinese PLA General Hospital, Beijing, 100010, People’s Republic of China, Email [email protected]
Background: While growing evidence suggests the intricate relationship between immune cells and the pathogenesis of acne, the causative implications underlying these associations remain poorly characterized. This study aimed to elucidate the causal links between various immune cell phenotypes and the development of acne using Mendelian randomization (MR) analysis.
Methods: Leveraging data of 731 immune cell traits and acne from genome-wide association studies, we conducted a bidirectional MR analysis. Rigorous instrumental variables selection was followed by causal inference using five complementary methods, including MR-Egger, weighted median, simple mode, inverse variance weighted (IVW), and weighted mode methods. Heterogeneity and pleiotropy were evaluated using Cochran’s Q test, MR-Egger intercept test, and leave-one-out analysis.
Results: Genetically predicted alterations in 26 immune cell phenotypes demonstrated causal associations with acne risk. Notably, 18 immune cell types exhibited protective effects, such as CD25 on IgD+ (IVW: OR 0.922, 95% CI 0.868– 0.979; p = 0.008), naive-mature B cell %lymphocytes (IVW: OR 0.824, 95% CI 0.698– 0.972; p = 0.022), and CD19 on sw mem (IVW: OR 0.841, 95% CI 0.752– 0.940; p = 0.002). Conversely, 8 immune cell types conferred increased risk, such as IgD+ CD38dim AC (IVW: OR 1.054, 95% CI 1.002– 1.108; p = 0.043), CD25 on unsw mem (IVW: OR 1.058, 95% CI 1.005– 1.114; p = 0.030), and CD28+ DN (CD4-CD8-) %DN (IVW: OR 1.117, 95% CI 1.019– 1.225; p = 0.019). The absence of significant heterogeneity or horizontal pleiotropy (p > 0.05) strengthens the credibility to the observed associations.
Conclusion: In conclusion, this research provides compelling genetic evidence for causal immunomodulatory influences on acne development, thus laying the groundwork for future investigational efforts aimed at uncovering targeted therapeutic strategies in acne management.
Keywords: circulating immune cells, acne, Mendelian randomization, genome wide association study
Introduction
Acne vulgaris, often known simply as acne, is a prevalent dermatological disorder distinguished by the inflammation of the pilosebaceous units. It typically emerges during adolescence and young adulthood, affecting an estimated 9.4% of individuals globally, and is recognized as the eighth most widespread illness around the world.1 This condition extends beyond mere cosmetic concerns; it can have a profound impact on a person’s self-worth and psychological well-being, potentially resulting in mental health issues, such as anxiety and depression.2 Moreover, the treatment and management of acne impose a considerable economic burden on both society and healthcare systems. Current therapeutic strategies for acne include topical medications, such as retinoids, benzoyl peroxide, and antibiotic ointments, in addition to oral retinoids, anti-androgen and antibiotics medications.3,4 Although these therapies demonstrate significant efficacy, they frequently present a range of adverse effects such as skin irritation, dryness, and alterations to the skin’s microecological balance. Furthermore, there is a growing apprehension surrounding the rise of bacteria that exhibit resistance to antibiotics.5 These limitations underscore the necessity for a more profound understanding of acne pathogenesis and the identification of novel therapeutic targets.
The development of acne involves a multifaceted interplay of various elements, including inflammatory reactions, excessive production of sebum, irregular keratinization within hair follicles, colonization by Cutibacterium acnes (C. acnes), and imbalanced secretion of androgens.6,7 The inflammatory reactions is a key characteristic of acne, influenced by various immunological factors. Recent research highlights the crucial roles of immune cells, including T cells, macrophages, and dendritic cells (DCs), in the development of acne.5,8 It is important to understand how these immune cells interact with the skin’s microbiome and contribute to inflammation and tissue damage, as this knowledge is vital for developing more effective and targeted acne treatments.
Mendelian randomization (MR) is an analytical approach that leverages genetic data, primarily used in epidemiology, to explore causal relationships in diseases. The goal of MR research is to assess whether individuals with certain genetic variations are at a higher risk of developing diseases compared to those without these variations, effectively “randomizing” genetic variants that affect risk factors. By utilizing genetic information, this method helps to clarify causal relationships in epidemiology, offering valuable insights into the underlying causes of diseases based on naturally occurring genetic differences.9 Recent applications of MR have yielded important findings in acne. A bidirectional MR analysis revealed 14 serum metabolites exhibiting significantly association with acne vulgaris.10 Another MR analysis indicated a mild protective effect of acne against schizophrenia and no effect of acne on the risk of depression, anxiety, obsessive-compulsive disorder, bipolar disorder and post-traumatic stress disorder.11 Li et al reported that there is no causal relationship between body mass index and acne using MR analysis.12
The two-sample MR analysis can utilize single-nucleotide polymorphism (SNP)-exposure and SNP-outcome associations derived from independent genome-wide association studies (GWASs) and combine them into a single causal estimate. This methodology benefits substantially from large-scale GWAS summary statistics, which enhance statistical power by increasing sample size and precision. In our study, we utilized two-sample MR to investigate the impact of immune cell characteristics on the onset of acne by taking advantage of genetic diversity while carefully excluding other potential confounding factors. We also performed reverse MR analysis, utilizing acne as the exposure variable and various immune cell characteristics as the outcome measures. Our results indicate that targeted modulation of specific immune cell populations could represent a promising strategy for the prevention or management of acne, though definitive causal evidence requires further research.
Materials and Methods
Study Design
This study utilized two-sample MR to explore the causal associations between 731 immune cell traits and acne. For a valid MR study, three essential conditions must be met. First, the genetic variants chosen as instrumental variables (IVs) are significant correlated with the exposure being examined, which is evaluated using the F-statistic. Typically, an F-statistic exceeding the value of 10 is deemed essential in order to mitigate the potential for weak instrument bias. Second, it is crucial that there are no unobserved confounding variables that could influence the relationship between the genetic variants and the outcome. Last, the genetic variations ought to influence the outcome solely via their effect on the exposure of interest, thereby avoiding horizontal pleiotropy. The research design is illustrated in Figure 1.
 |
Figure 1 (A) The diagram of MR assumption. (B) The diagram of MR analysis processing. Abbreviations: SNPs, single nucleotide polymorphisms; GWAS, genome-wide association study; MR, Mendelian randomization; MR-PRESSO, Mendelian Randomization Pleiotropy RESidual Sum and Outlier (MR-PRESSO) test. |
Data Source for the Immune Cells and Acne
The data of GWAS encompassed 731 immune cell phenotypes collected from a cohort consisting of 3757 individuals of European descent residing in Sardinia, Italy.13 To identify genetic variants linked to these immune cells, researchers utilized a reference panel consisting of 3514 Sardinian individuals and analyzed SNPs with high-density arrays technology.13 The data underwent adjustments for factors such as sex, age, and the square of age. Initially, the immune cell populations were categorized based on seven major flow cytometry panels, which included the B cell panel, T-cell maturation stages panel, conventional DC (cDC) panel, Monocyte panel, Myeloid cell panel, T lymphocytes, B lymphocytes, Natural Killer cells (TBNK) panel, and Regulatory T cell (Treg) panel.
The GWAS data for acne included a substantial cohort of 212,334 participants of European ancestry, with 1299 individuals with acne and 211,139 individuals serving as controls. Following rigorous quality control measures and estimation processes, the analysis focused on 16,380,454 SNPs. For those interested in further details, the complete dataset can be accessed through the Finn-B-L12 acne dataset (https://gwas.mrcieu.ac.uk/datasets/finn-b-L12_acne/).
Selection of IVs
Given that too few IVs were obtained when using the strict significance threshold (p < 5 × 10−8) as the selection criterion for IVs, we relaxed the significance threshold to p < 1 × 10−5.10,14 To reduce the impact of weak instrument bias, SNPs are evaluated using the F-statistic. Additionally, SNPs in linkage disequilibrium are excluded, specifically those with an r² value lower than 0.001 and a spatial separation exceeding than 10,000 kilobases, to ensure their independence.15
Statistical Analysis
All analyses were performed utilizing R (version 4.3.0) and incorporated five MR approaches, specifically MR-Egger, simple mode, weighted median (WM), inverse variance weighted (IVW), and weighted mode, to assess the causal relationships between immune cell variations and the risk of developing acne. The main outcome was obtained from the IVW method, which indicated a statistically significant finding with a p-value below 0.05. The IVW method is advantageous as it emphasizes less variable variants, leading to more accurate estimates.9 In contrast, the WM method stipulates that a minimum of 50% of the weight utilized in the analysis must originate from valid instruments to ensure reliable causal estimates.16 Furthermore, we evaluated heterogeneity through Cochran’s Q statistic, which assesses the consistency of genetic variation effects on phenotypes. A significant Cochran’s Q statistic, indicated p-value lower than 0.05, points to considerable variability in the effects of different genetic variations.17 The MR-Egger method was employed to assess horizontal pleiotropy.16 The identification of outlier IVs and the adjustment for horizontal pleiotropy were accomplished through the utilization of the MR-Pleiotropy Residual Sum and Outlier (MR-PRESSO) methodology.
We utilized the same MR methods to investigate the potential impact of acne on immune cell traits, aiming to mitigate issues of reverse causality and confounding that often arise in epidemiological studies. In this reverse MR analysis, acne was designated as the exposure variable, while various immune cell characteristics were assessed as the outcome measures.
Results
In order to evaluate the causal impacts of different immune cell types on acne, we conducted a two-sample MR analysis, while also considering potential confounders and the possibility of horizontal pleiotropy. Our analysis identified 26 immune cell types that are significantly associated with acne, with a statistical significance threshold set at p < 1 × 10−5 (Figure 2 and Table S1). The robustness of our findings was further confirmed through scatter plots and the “leave-one-out” approach, which are presented in Figures S1 and S2.
 |
Figure 2 Forest plot shows the effect of immune cells on acne. |
Within B cell subsets, three characteristics were identified as positively correlated with acne risk: IgD+ CD38dim AC [(IVW: odds ratio (OR) 1.054, 95% confidence interval (CI) 1.002–1.108; p = 0.043)], CD25 on unsw mem (IVW: OR 1.058, 95% CI 1.005–1.114; p = 0.030), and CD24 on sw mem (IVW: OR 1.091, 95% CI 1.017–1.170; p = 0.015). Conversely, four traits exhibited protective effect on acne: CD25 on IgD+ (IVW: OR 0.922, 95% CI 0.868–0.979; p = 0.008), Naive-mature B cell %lymphocyte (IVW: OR 0.824, 95% CI 0.698–0.972; p = 0.022), CD19 on sw mem (IVW: OR 0.841, 95% CI 0.752–0.940; p = 0.002), and PB/PC AC (IVW: OR 0.921, 95% CI 0.866–0.981; p = 0.010).
Surprisingly, all four significant traits within the TBNK subsets were protective: DN (CD4-CD8-) %T cell (IVW: OR 0.838, 95% CI 0.710–0.989; p = 0.037), CD8dim NKT AC (IVW: OR 0.912, 95% CI 0.837–0.994; p = 0.035), B cell %lymphocyte (IVW: OR 0.904, 95% CI 0.824–0.992; p = 0.032), and DN (CD4-CD8-) AC (IVW: OR 0.822, 95% CI 0.715–0.946; p = 0.006).
Within cDC subsets, two traits were linked with an increased risk of acne: SSC-A on myeloid DC (IVW: OR 1.059, 95% CI 1.003–1.119; p = 0.039), and CD62L- myeloid DC AC (IVW: OR 1.176, 95% CI 1.055–1.311; p = 0.003).
Within the Treg cell subsets, one trait was linked with increased risk of acne: CD28+ DN (CD4-CD8-) %DN (IVW: OR 1.117, 95% CI 1.019–1.225; p = 0.019). Conversely, eight traits demonstrated inverse associations with acne risk: CD28- DN (CD4-CD8-) %T cell (IVW: OR 0.843, 95% CI 0.762–0.933; p = 0.001), CD25 on CD39+ secreting Treg (IVW: OR 0.929, 95% CI 0.873–0.989; p = 0.021), CD39+ secreting Treg %secreting Treg (IVW: OR 0.936, 95% CI 0.878–0.997; p = 0.040), CD39+ secreting Treg%CD4 Treg (IVW: OR 0.930, 95% CI 0.876–0.988; p = 0.018), CD28 on CD39+ activated Treg (IVW: OR 0.888, 95% CI 0.825–0.956; p = 0.002), Secreting Treg AC (IVW: OR 0.962, 95% CI 0.928–0.998; p = 0.040), CD28- DN (CD4-CD8-) %DN (IVW: OR 0.895, 95% CI 0.817–0.982; p = 0.019), and Activated & secreting Treg AC (IVW: OR 0.962, 95% CI 0.928–0.996; p = 0.029).
Within myeloid cell panels, two traits were identified as being associated with decreased risk of acne: CD11b on Mo MDSC (IVW: OR 0.921, 95% CI 0.858–0.989; p = 0.024), and HLA DR on CD33- HLA DR+ (IVW: OR 0.924, 95% CI 0.859–0.995; p = 0.037).
Two traits within the maturation stages of T-cell subsets were associated with increased risk of acne: CD8 on CM CD8br (IVW: OR 1.229, 95% CI 1.059–1.427; p = 0.007), and CM DN (CD4-CD8-) %T cell (IVW: OR 1.128, 95% CI 1.003–1.269; p = 0.045).
In addition, we employed a reverse MR approach to explore potential reverse causation between acne and immune cell traits, where acne may influence the proportion and phenotype of immune cells (Figure 3 and Table S2). The results of this analysis provide intriguing evidence of an association between acne and specific immune cell populations.
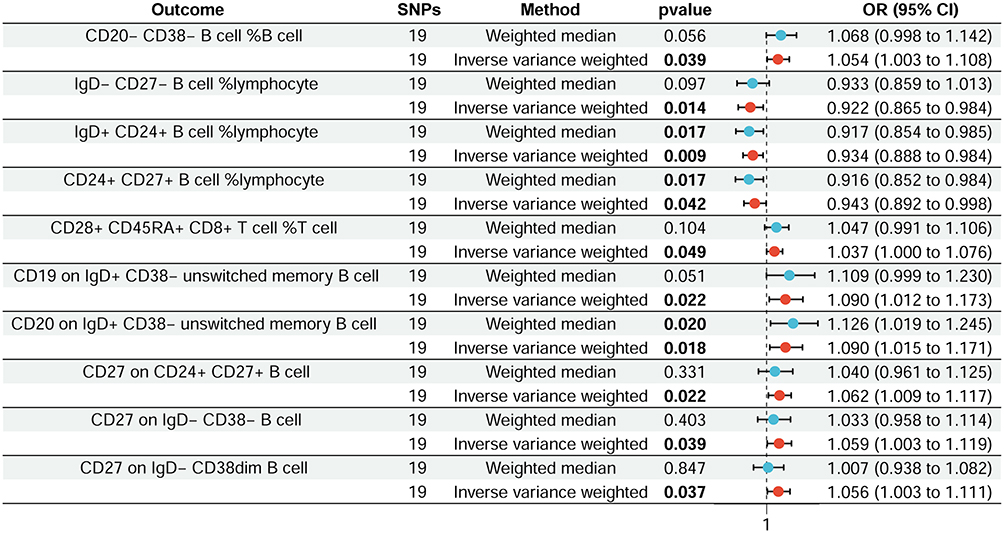 |
Figure 3 Forest plot shows the effect of acne on immune cells. |
The results indicate positive links between acne and CD20- CD38- B cell %B cell (IVW: OR 1.054, 95% CI 1.003–1.108; p = 0.039), CD28+ CD45RA+ CD8+ T cell %T cell (IVW: OR 1.037, 95% CI 1.000–1.076; p = 0.049), CD19 on IgD+ CD38- unswitched memory B cell (IVW: OR 1.090, 95% CI 1.012–1.173; p = 0.022), CD20 on IgD+ CD38- unswitched memory B cell (IVW: OR 1.090, 95% CI 1.015–1.171; p = 0.018), CD27 on CD24+ CD27+ B cell (IVW: OR 1.062, 95% CI 1.009–1.117; p = 0.022), CD27 on IgD- CD38- B cell (IVW: OR 1.059, 95% CI 1.003–1.119; p = 0.039), and CD27 on IgD- CD38dim B cell (IVW: OR 1.0560, 95% CI 1.003–1.111; p = 0.037). Conversely, negative correlations were observed for IgD- CD27- B cell%lymphocyte (IVW: OR 0.922, 95% CI 0.865–0.984; p = 0.014), IgD+ CD24+ B cell%lymphocyte (IVW: OR 0.934, 95% CI 0.888–0.984; p = 0.009), and CD24+ CD27+ B cell%lymphocyte (IVW: OR 0.944, 95% CI 0.892–0.998; p = 0.043). Further research is warranted to clarify the mechanisms behind these associations and their implications for acne treatment.
Our sensitivity analysis corroborated the findings, confirming the robustness of our results through heterogeneity and pleiotropy assessments (Tables 1 and 2). Leave-one-out cross-validation further substantiated these conclusions.
 |
Table 1 Pleiotropy and Heterogeneity Test of the MR Analysis |
 |
Table 2 Pleiotropy and Heterogeneity Test of the Reverse MR Analysis |
Discussion
In the present investigation, we utilized MR analysis to mitigate confounding and reverse causation biases to examine the casual links between acne and immune cell traits. We discovered multiple notable characteristics of immune cells were linked to the risk of developing acne, with a dichotomy of 8 traits indicating increased risk and 18 showing protective effects. In the reverse MR analysis, we also observed some positive results. These findings not only enhance our knowledge of the immunological contributors to acne but also hold promise for the development of targeted immunomodulatory therapies, potentially transforming acne management and alleviating the burden on affected individuals.
The interplay between B lymphocytes and acne vulgaris has emerged as a burgeoning area of interest. The B lymphocytes can promote the activity of other immune cells by producing specific cytokines, thereby exacerbating inflammation. By examining a series of cell surface markers in lymphocytes, it was found that an increase in B cells was positively correlated with the severity of acne, indicating that B cells play an indispensable role in the immune response of acne.18 In acne, B lymphocytes had been implicated in the formation of long-standing scars. Gene expression and immunohistochemistry analyses showed B-cell infiltrations were not typically found in early papules of acne but were involved in 23% of all acne scar specimens.19,20 Although the production of antibodies is a hallmark of B cells, it is increasingly recognized that B cell dysfunction can contribute to the production of inflammatory cytokines. This suggests that, in addition to their well-known role in humoral immunity, B cells may also play a significant role in the inflammatory processes associated with acne pathogenesis. Our study highlighted that specific B cell subsets, namely IgD+ CD38dim AC, CD25 on unsw mem, and CD24 on sw mem, are implicated in the increased risk of acne development. In contrast, other subsets, including CD25 on IgD+, Naive-mature B cell %lymphocyte, CD19 on sw mem, and PB/PC AC, appear to confer a protective effect against the disease. These observations may prompt an exploration into the roles of CD38, CD24, CD25, and CD19 in the pathogenesis of acne, potentially elucidating their immunomodulatory functions within the complex cytokine milieu of acne lesions. The intricate balance between these B cell markers and their influence on the immune response may offer novel pathways for treatment strategies.
The exploration of TBNK panels in the context of acne vulgaris has unveiled intriguing connections between specific lymphocyte subsets and the disease’s pathogenesis. Recent studies have shed light on the potential protective roles of certain T and B cell populations.21,22 Flow cytometric analysis indicated that individuals with severe acne exhibited a notable increase in NK cell levels compared to those with moderate acne, indicating a potential role of NK cells in the development of acne.23 Additionally, the infiltration level of NK cell in the acne lesions implies their participation in localized immune reactions.24 Our research corroborates these findings, demonstrating that all four significant traits within TBNK panels exert a protective effect: DN (CD4-CD8-) %T cell, CD8dim NKT AC, B cell %lymphocyte, and DN (CD4-CD8-) AC.
DCs bridge innate and adaptive immunity by sensing and presenting antigens to initiate immune responses.25 They can be classified into four types through unsupervised analysis of conventional flow cytometry and mass cytometry data, including Langerhans cells (LCs), cDCs, plasmacytoid DCs (pDCs), as well as DCs derived from monocytes.26 Skin immunohistochemistry revealed significantly increased numbers of CD1+ (LCs) and CD83+ DCs in early acne than nonlesional skin, suggesting they play a role in lesion development.27,28 LCs may present C. acnes antigens to CD4+ T cells after environmental changes boost their production.29 Activated DCs produce IL-1 and IL-23, key in Th17 cell activation.30 Studies have discovered that the levels of cytokines in the lesions of acne patients were markedly increased, further propelling the worsening of local inflammation. During the early stages of acne, the activation of DCs can promote the infiltration of T cells, especially Th1 and Th17 cells. These cells release a multitude of pro-inflammatory cytokines, resulting in exacerbated inflammation in the skin tissue and the development of typical acne lesions.31 “SSC-A on myeloid DC (cDC panel)” measures the side scatter signal intensity in myeloid DCs within the cDC subset. SSC-A reflects cell size, shape, and complexity as detected by flow cytometry. Higher SSC-A suggests a larger cell population but does not count actual cells.14 These findings implicate the important role of DCs, particularly of the myeloid lineage, in both the onset and development of acne lesions. The upregulation of CD62L expression on myeloid DCs likely reflect their active role in sculpting the typical inflammatory environment of acne. Further research into the functional alterations of these cDC subsets and their interactions with other immune cells will be critical to elucidate their role in acne pathogenesis fully.
In the early stages of acne lesions, a significantly higher numbers of Foxp3+ cells had been observed in the dermis of lesional than non-lesional skin as assessed by immunohistochemistry.27,32 Tregs in individuals with acne may have a functional deficiency, which prevents them from suppressing the persistent immune response seen in acne lesions effectively. Under inflammatory conditions, Tregs can forfeit their inhibitory capabilities and transform into cells that express interleukin-17 (IL-17), potentially contributing to chronic skin inflammation.33,34 Furthermore, the count of Tregs is noted to be lower in the affected skin of acne patients compared to unaffected skin by integrated bioinformatics analysis.35 The immunopathogenesis of acne vulgaris might be associated with an imbalance in the Th17/Treg ratio, where an increased Th17/Treg ratio could initiate inflammatory processes and negatively impact the hair follicle’s homeostasis and integrity controlled by Tregs.36 Retinoids, known to be beneficial in acne treatment, are thought to work by inhibiting IL-17 and increasing Foxp3 expression, thus helping to modulate the equilibrium between Treg and Th17 cell differentiation.37 Our research contributes to this field by identifying specific Treg-associated traits associated with an elevated risk of acne: CD28+ within the double-negative (DN) subset (CD4-CD8-). In contrast, several traits have shown protective associations, such as CD28- DN (CD4-CD8-) T cells, CD25 on CD39+ secreting Tregs, and CD39+ secreting Tregs %secreting Tregs, suggesting a complex regulatory role for Tregs in acne immunopathogenesis. The identification of these Treg subsets in the context of acne suggests a delicate balance that could influence disease progression or resolution.
Myeloid cells, which encompass monocytes, macrophages, and DCs, play a multifaceted role in acne and other skin inflammatory diseases, not only participating in the clearance of pathogens and the regulation of inflammatory responses but also being involved in scar formation and treatment responses.31 In a study combining single-cell and spatial RNA sequencing with ultra-high resolution Seq-Scope analysis, TREM2 macrophages were identified near hair follicle epithelium expressing squalene epoxidase in early acne lesions, which were abundant in acne lesions, expressing genes related to lipid metabolism and inflammation. The research found that squalene, a lipid found in high concentrations in acne lesions, induced the differentiation of TREM2 macrophages in vitro that were unable to kill C. acnes due to squalene’s inhibitory effect on oxidative enzymes and oxygen free radical scavenging.38 Our research indicates that within myeloid cell subsets, CD11b expression on Mo MDSC and HLA-DR expression on CD33- HLA DR+ cells are linked to protective effects against acne development. The observed protective associations may indicate the cells’ ability to mitigate excessive inflammation, a concept supported by research over the past decade highlighting the immunomodulatory functions of myeloid cells in skin health.21,24 Elucidating the interplay between myeloid cells and acne may facilitate the creation of innovative treatment approaches that utilize their regulatory functions.
In early acne lesions, CD4+ T helper cells, particularly Th1 and Th17 subsets, are predominantly infiltrated. These cells are crucial in the development of acne and they participate in the immune response that triggers inflammation. They contribute to the inflammatory process by producing cytokines, such as IL-17, which further attracts other immune cells, including eosinophils and neutrophils, thereby exacerbating skin inflammation.39 Our study contributes to this field by identifying a specific trait within T-cell maturation stages that is associated with an increased risk of acne: the expression of CD8 on central memory (CM) CD8+ (CM CD8br) and double-negative (DN) T cells (CD4-CD8-). This finding suggests that specific stages of T-cell maturation may play a critical role in acne development, potentially modulating the adaptive immune response and the skin’s inflammatory processes.
Our findings that specific immune cell phenotype causally influence acne risk align with emerging therapeutic strategies targeting immunomodulatory pathways in dermatology. Amlitelimab, a non-depleting anti-OX40 ligand monoclonal antibody, demonstrated sustained efficacy in atopic dermatitis by modulating T-cell-driven inflammation (phase 2b study).40 Xeligekimab (GR1501), a fully human monoclonal antibody that selectively neutralizes IL-17A and showed high efficacy and was well tolerated in Chinese patients with moderate-to-severe plaque psoriasis (Phase III study).41 These parallels underscore the translational potential of our results, suggesting that targeting specific immune cell phenotypes may offer analogous therapeutic avenues for acne management.
We must acknowledge certain inherent limitations in our study. Firstly, we employed a threshold of p < 10−5 for IVs selection due to limited exposure sizes. Lowering the p-value threshold might influence the statistical power of the overall MR analysis. Secondly, the datasets for immune traits and acne were derived from different ethnic groups. Although extensive sensitivity analyses were performed to mitigate confounding factors, some racial heterogeneity persists in this MR analysis. Thirdly, the immune trait and acne datasets are at the summary level, lacking individual-level data. Consequently, stratification analyses specific to acne patients are not feasible.
Conclusion
In conclusion, this study elucidate intricate causal interaction patterns among various immune traits and acne through MR analysis, underscoring the significant impact of immune cells in driving acne vulgaris progression. Our findings indicated 18 immune cell types associated with decreased risk of acne, such as CD25 on IgD+, naive-mature B cell %lymphocytes, and CD19 on sw mem, and 8 immune cell types correlated with increased risk of acne, such as IgD+ CD38dim AC, CD25 on unsw mem, and CD28+ DN (CD4-CD8-) %DN. The identification of specific immune cell subsets and their phenotypic markers lays the groundwork for targeted therapies that might modulate these immunological pathways. As our understanding of acne’s immunological landscape deepens, we move closer to developing personalized treatments that address both symptoms and the underlying causes.
Data Sharing Statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics Approval and Informed Consent
The Ethical Committee of Seventh Medical Center of Chinese PLA General Hospital (S2024-072-01) approved this study and waived the requirement for written informed consent for all data are taken from public databases and are exempt from ethical review.
Acknowledgments
We gratefully thank the European Bioinformatics Institute for providing statistical data.
Author Contributions
All authors (X.W., S.G., Z.X., X.Z., Y.B., Q.L., H.L. and R.Y.) significantly contributed to the study’s conception, design, data handling, and interpretation. They were involved in drafting and revising the manuscript, approved the final version, agreed on the target journal, and are accountable for the work’s integrity.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Disclosure
The authors declare that they have no conflicts of interest.
References
1. Tan JK, Bhate K. A global perspective on the epidemiology of acne. Br J Dermatol. 2015;172(Suppl 1):3–12. doi:10.1111/bjd.13462
2. Gieler U, Gieler T, Kupfer JP. Acne and quality of life - impact and management. J Eur Acad Dermatol Venereol. 2015;29(Suppl 4):12–14. doi:10.1111/jdv.13191
3. Oge LK, Broussard A, Marshall MD. Acne Vulgaris: diagnosis and Treatment. Am Fam Physician. 2019;100(8):475–484.
4. Leyden J, Stein-Gold L, Weiss J. Why topical retinoids are mainstay of therapy for acne. Dermatol Ther. 2017;7(3):293–304. doi:10.1007/s13555-017-0185-2
5. Dréno B, Dagnelie MA, Khammari A, Corvec S. The skin microbiome: a new actor in inflammatory acne. Am J Clin Dermatol. 2020;21(Suppl 1):18–24. doi:10.1007/s40257-020-00531-1
6. Williams HC, Dellavalle RP, Garner S. Acne vulgaris. Lancet. 2012;379(9813):361–372. doi:10.1016/S0140-6736(11)60321-8
7. Melnik BC. Acne transcriptomics: fundamentals of acne pathogenesis and isotretinoin treatment. Cells. 2023;12(22). doi:10.3390/cells12222600
8. Zouboulis CC, Jourdan E, Picardo M. Acne is an inflammatory disease and alterations of sebum composition initiate acne lesions. J Eur Acad Dermatol Venereol. 2014;28(5):527–532. doi:10.1111/jdv.12298
9. Burgess S, Butterworth A, Thompson SG. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet Epidemiol. 2013;37(7):658–665. doi:10.1002/gepi.21758
10. Wang X, Wu Y, Zhao P, Wang X, Wu W, Yang J. The causal relationship between serum metabolites and acne vulgaris: a Mendelian randomization study. Sci Rep. 2024;14(1):11045. doi:10.1038/s41598-024-61850-5
11. Liu L, Xue Y, Chen Y, et al. Acne and risk of mental disorders: a two-sample Mendelian randomization study based on large genome-wide association data. Front Public Health. 2023;11:1156522. doi:10.3389/fpubh.2023.1156522
12. Li Z, Qi W, Zang T, Zhang Z. The causal relationship between acne vulgaris and BMI: a Mendelian randomization study. J Cosmet Dermatol. 2025;24(3):e70092. doi:10.1111/jocd.70092
13. Orrù V, Steri M, Sidore C, et al. Complex genetic signatures in immune cells underlie autoimmunity and inform therapy. Nat Genet. 2020;52(10):1036–1045. doi:10.1038/s41588-020-0684-4
14. Wang C, Zhu D, Zhang D, et al. Causal role of immune cells in schizophrenia: Mendelian randomization (MR) study. BMC Psychiatry. 2023;23(1):590. doi:10.1186/s12888-023-05081-4
15. Hosier H, Lipkind HS, Rasheed H, DeWan AT, Rogne T. Dyslipidemia and risk of preeclampsia: a multiancestry Mendelian randomization study. Hypertension. 2023;80(5):1067–1076. doi:10.1161/HYPERTENSIONAHA.122.20426
16. Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur J Epidemiol. 2017;32(5):377–389. doi:10.1007/s10654-017-0255-x
17. Cohen JF, Chalumeau M, Cohen R, Korevaar DA, Khoshnood B, Bossuyt PM. Cochran’s Q test was useful to assess heterogeneity in likelihood ratios in studies of diagnostic accuracy. J Clin Epidemiol. 2015;68(3):299–306. doi:10.1016/j.jclinepi.2014.09.005
18. Holland DB, Gowland G, Cunliffe WJ. Lymphocyte subpopulations in patients with acne vulgaris. Br J Dermatol. 1983;109(2):199–203. doi:10.1111/j.1365-2133.1983.tb07081.x
19. Carlavan I, Bertino B, Rivier M, et al. Atrophic scar formation in patients with acne involves long-acting immune responses with plasma cells and alteration of sebaceous glands. Br J Dermatol. 2018;179(4):906–917. doi:10.1111/bjd.16680
20. Chancheewa B, Asawanonda P, Noppakun N, Kumtornrut C. Myofibroblasts, B cells, and mast cells in different types of long-standing acne scars. Skin Appendage Disord. 2022;8(6):469–475. doi:10.1159/000524566
21. Huang L, Yang S, Yu X, et al. Association of different cell types and inflammation in early acne vulgaris. Front Immunol. 2024;15:1275269. doi:10.3389/fimmu.2024.1275269
22. Jin Z, Song Y, He L. A review of skin immune processes in acne. Front Immunol. 2023;14:1324930. doi:10.3389/fimmu.2023.1324930
23. Gunaydin SD, Tezcan I. Evaluation of peripheral lymphocyte subsets in acne vulgaris patients before and after systemic isotretinoin treatment. Indian J Pharmacol. 2022;54(5):338–344. doi:10.4103/ijp.ijp_695_21
24. Firlej E, Kowalska W, Szymaszek K, Roliński J, Bartosińska J. The Role of Skin Immune System in Acne. J Clin Med. 2022;11(6):1579. doi:10.3390/jcm11061579
25. Dress RJ, Wong AY, Ginhoux F. Homeostatic control of dendritic cell numbers and differentiation. Immunol cell biol. 2018;96(5):463–476. doi:10.1111/imcb.12028
26. Guilliams M, Dutertre CA, Scott CL, et al. Unsupervised high-dimensional analysis aligns dendritic cells across tissues and species. Immunity. 2016;45(3):669–684. doi:10.1016/j.immuni.2016.08.015
27. Kelhälä HL, Palatsi R, Fyhrquist N, et al. IL-17/Th17 pathway is activated in acne lesions. PLoS One. 2014;9(8):e105238. doi:10.1371/journal.pone.0105238
28. Layton AM, Morris C, Cunliffe WJ, Ingham E. Immunohistochemical investigation of evolving inflammation in lesions of acne vulgaris. Exp Dermatol. 1998;7(4):191–197. doi:10.1111/j.1600-0625.1998.tb00323.x
29. Farrar MD, Ingham E. Acne: inflammation. Clin Dermatol. 2004;22(5):380–384. doi:10.1016/j.clindermatol.2004.03.006
30. van Beelen AJ, Zelinkova Z, Taanman-Kueter EW, et al. Stimulation of the intracellular bacterial sensor NOD2 programs dendritic cells to promote interleukin-17 production in human memory T cells. Immunity. 2007;27(4):660–669. doi:10.1016/j.immuni.2007.08.013
31. Feng Y, Li J, Mo X, Ju Q. Macrophages in acne vulgaris: mediating phagocytosis, inflammation, scar formation, and therapeutic implications. Front Immunol. 2024;15:1355455. doi:10.3389/fimmu.2024.1355455
32. Farag AGA, Maraee AH, Rifaat Al‐Sharaky D, Elshaib ME, Kohla MSM, Shehata WA. Tissue expression of IL‐17A and FOXP3 in acne vulgaris patients. J Cosmet Dermatol. 2020;20(1):330–337. doi:10.1111/jocd.13485
33. Bovenschen HJ, van de Kerkhof PC, van Erp PE, Woestenenk R, Joosten I, Koenen HJ. Foxp3+ regulatory T cells of psoriasis patients easily differentiate into IL-17A-producing cells and are found in lesional skin. J Invest Dermatol. 2011;131(9):1853–1860. doi:10.1038/jid.2011.139
34. Jung MK, Kwak JE, Shin EC. IL-17A-producing Foxp3(+) regulatory T cells and human diseases. Immune Netw. 2017;17(5):276–286. doi:10.4110/in.2017.17.5.276
35. Yang L, Shou YH, Yang YS, Xu JH. Elucidating the immune infiltration in acne and its comparison with rosacea by integrated bioinformatics analysis. PLoS One. 2021;16(3):e0248650. doi:10.1371/journal.pone.0248650
36. Melnik BC, John SM, Chen W, Plewig G. T helper 17 cell/regulatory T-cell imbalance in hidradenitis suppurativa/acne inversa: the link to hair follicle dissection, obesity, smoking and autoimmune comorbidities. Br J Dermatol. 2018;179(2):260–272. doi:10.1111/bjd.16561
37. Elias KM, Laurence A, Davidson TS, et al. Retinoic acid inhibits Th17 polarization and enhances FoxP3 expression through a Stat-3/Stat-5 independent signaling pathway. Blood. 2008;111(3):1013–1020. doi:10.1182/blood-2007-06-096438
38. Do TH, Ma F, Andrade PR, et al. TREM2 macrophages induced by human lipids drive inflammation in acne lesions. Sci Immunol. 2022;7(73):eabo2787. doi:10.1126/sciimmunol.abo2787
39. Mias C, Mengeaud V, Bessou-Touya S, Duplan H. Recent advances in understanding inflammatory acne: deciphering the relationship between Cutibacterium acnes and Th17 inflammatory pathway. J Eur Acad Dermatol Venereol. 2023;37(Suppl 2):3–11. doi:10.1111/jdv.18794
40. Weidinger S, Blauvelt A, Papp KA, et al. Phase 2b randomized clinical trial of amlitelimab, an anti-OX40 ligand antibody, in patients with moderate-to-severe atopic dermatitis. J Allergy Clin Immunol. 2025;155(4):1264–1275. doi:10.1016/j.jaci.2024.10.031
41. Cai L, Jiang C, Zhang G, et al. A multicentre randomized double-blind placebo-controlled phase III study of the efficacy and safety of xeligekimab (GR1501) in patients with moderate-to-severe plaque psoriasis. Br J Dermatol. 2024;191(3):336–343. doi:10.1093/bjd/ljae062
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Revealing the Impact of Gut Microbiota on Acne Through Mendelian Randomization Analysis

Ji X, Wu S, Zhao D, Bai Q, Wang Y, Gong K, Zheng H, Zhu M
Clinical, Cosmetic and Investigational Dermatology 2024, 17:383-393
Published Date: 8 February 2024