")
Back to Journals » Journal of Inflammation Research » Volume 17
New Posttranslational Modification Lactylation Brings New Inspiration for the Treatment of Rheumatoid Arthritis
Authors Yang Y , Shi J, Yu J, Zhao X, Zhu K, Wang S, Zhang X, Zhang X, Wei G , Cao W
Received 21 September 2024
Accepted for publication 21 December 2024
Published 30 December 2024 Volume 2024:17 Pages 11845—11860
DOI https://doi.org/10.2147/JIR.S497240
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Yue Yang,1,* Jinjie Shi,2,* Jiming Yu,3,* Xin Zhao,1 Ke Zhu,1 Shen Wang,4 Xinwen Zhang,1 Xieyu Zhang,1 Guangcheng Wei,1 Wei Cao1
1Rheumatology Department, Wangjing Hospital of China Academy of Chinese Medical Sciences, Beijing, People’s Republic of China; 2Graduate School, Beijing University of Chinese Medicine, Beijing, People’s Republic of China; 3The Second Clinical Medical College, Henan University of Chinese Medicine, Zhengzhou, People’s Republic of China; 4Orthopedics Department, Nanjing Integrated Traditional Chinese and Western Medicine Hospital, Nanjing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Wei Cao, Wangjing Hospital of China Academy of Chinese Medical Sciences, 6 South Central Road, Chaoyang District, Beijing, 100102, People’s Republic of China, Email [email protected]
Abstract: Lactic acid (LA) is an essential glycolytic metabolite and energy source in the body, which is present in high levels in the synovial fluid of patients with rheumatoid arthritis (RA) and is a reliable indicator for identifying inflammatory arthritis. LA not only acts as an inflammatory amplifier in RA, recent studies have found that novel posttranslational modification (PTM) lactylation mediated by LA may also play a key role in RA. Single-cell sequencing showed that the RA lactylation score of patients with RA was significantly increased, and core lactylation-promoting genes, including NDUFB3, NGLY1, and other genes, were found to be potential biomarkers of RA. More studies have shown that lactylation can regulate genes in various cells, such as fibroblast-like synoviocytes (FLSs) and macrophages, thus playing a special role in the development and occurrence of autoimmune diseases, neurological diseases, and cancer diseases. In this paper, we review the research on lactylation in RA-related cells and mechanisms and bring new insights into the pathogenesis, diagnosis, and treatment of RA.
Keywords: lactic acid, lactylation, posttranslational modification, rheumatoid arthritis
Introduction
Rheumatoid arthritis (RA) is a persistent and inflammatory joint disease caused by the immune system attacking immunoglobulin G (IgG) and citrullinated proteins. These attacks are identified by the presence of autoantibodies known as rheumatoid factor (RF) and anti-citrullinated protein antibodies (ACPAs). The development of RA is influenced by genetic, epigenetic, and environmental factors.1 The majority of epidemiological investigations on RA have been conducted in Western countries. These studies have revealed a prevalence of RA ranging from 0.5% to 1.0% among individuals of white ethnicity.2,3 If not addressed, these conditions can manifest as small areas of tissue death, the sticking together of granulation tissue, and the formation of fibrous tissue on the joint’s surface. This can result in the gradual stiffening of the joint, its deterioration, deformities, and functional impairment.4 Furthermore, at the advanced stage of RA, there is a heightened risk of developing respiratory, circulatory, and tumour-related illnesses, which significantly increases the likelihood of mortality. RA’s prevalence and significant ramifications have garnered considerable interest, leading to a growing study on the subject.5 Currently, the precise cause of RA is unknown. Although numerous drugs, such as nonsteroidal anti-inflammatory drugs (NSAIDs), corticosteroids, and disease-modifying antirheumatic drugs (DMARDs), have been used, it remains undeniable that a significant number of patients may encounter adverse reactions and treatment failure, including nonresponse and limited effectiveness.6,7 Therefore, it is necessary to continue exploring RA’s critical pathogenesis.
 |
Figure 1 The process of lactylation. As a substrate for lactylation, the metabolite LA can enter the nucleus to bind to histone lysine residues or bind to non-histone lysine residues in vivo to form protein modifications. In turn, it regulates gene expression and affects cellular activity. |
 |
Figure 2 Network of relationships among PTMs, lactylation, LA, and RA. A variety of PTMs are related to the occurrence and development of RA. As a metabolite of glycolysis, LA exists in the synovium of the hypoxic microenvironment, which affects the progression of RA in many ways. LA accumulation was found to cause protein lactylation modification, a type of PTM, and may affect RA through critical cells and essential mechanisms of RA. |
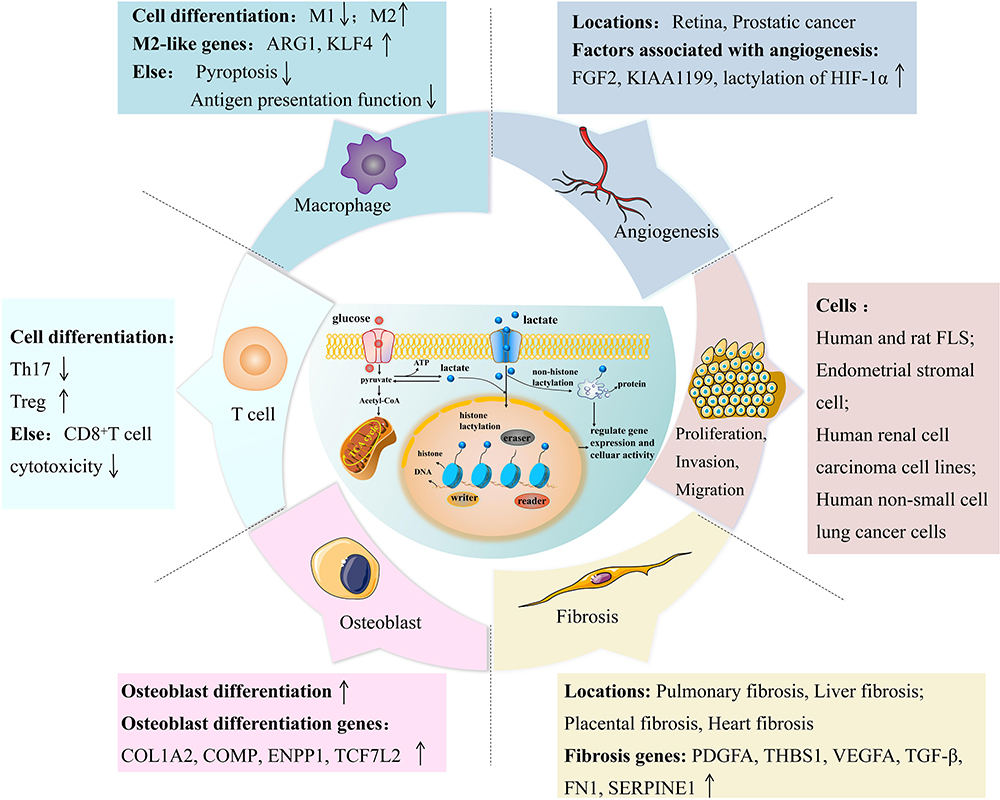 |
Figure 3 Lactylation and critical cells and essential mechanisms of RA. (1) Lactylation modification can promote the process of converting pro-inflammatory M1 macrophages into anti-inflammatory M2 macrophages and promote the expression of M2-like genes Arg1 and Klf4. Moreover, it can inhibit the pyroptosis and antigen presentation of macrophages. (2) On the T cell side, lactylation reprograms proinflammatory Th17 cells into regulatory T cells and may additionally attenuate CD8+ T cell effector function. (3) Lactylation can promote osteogenic differentiation and enhance the expression of osteogenic genes. (4) Lactylation can enhance the expression of pro-angiogenic factors such as FGF2 and promote angiogenesis in retina and prostate cancer. (5) Lactylation can promote the proliferation, invasion and migration of human and rat FLS, endometrial stromal cells, human renal cell carcinoma cell lines, and human non-small cell lung cancer cells. (6) Lactylation can also regulate fibrosis genes such as PDGFA, affecting pulmonary, liver, placental, and heart fibrosis. |
Recent studies have demonstrated that epigenetic regulators, including histone modifications, methylation of DNA and RNA, and non-coding RNAs, are crucial in the development of RA.8 Moreover, current research has discovered that certain metabolites within the body serve as signalling molecules in the regulation of genes. One method by which these metabolites communicate is by the chemical alteration of proteins, such as histones. It is classified as a posttranslational modification (PTM). Zhang et al have identified a new histone modification called lactylation, produced from the biological metabolite lactic acid (LA).9,10 Moreover, in the synovial fluid of patients with RA, high levels of LA accumulate due to the high metabolic demand of synovial cells, making it a reliable indicator for differentiating inflammatory arthritis.11,12 In turn, the LA accumulated in RA synovium acts on fibroblast-like synoviocytes (FLSs), macrophages, T cells, and other cells, becoming an amplifier of inflammation.13 Studies also showed that LA can activate hypoxia-inducible factor-1α (HIF-1α) in bovine FLSs through the PI3K/Akt/ NF-κB signaling pathway to enhance the gene expression of pro-inflammatory factor interleukin-6 (IL-6).11 On the other hand, With the discovery of lactylation, more and more studies also have found that lactylation may play a key role in cancer, autoimmune diseases, and other diseases by regulating gene expression in various cells, such as macrophages and T cells.14 First discovered in macrophages, lactylation may induce macrophage M2 signature by initiating the expression of homeostasis genes.10 The latest single-cell sequencing study found that RA lactylation score was significantly increased in patients with RA, which was positively correlated with immune cell infiltration and immune checkpoint molecular expression. The study also confirmed that core lactylation-promoting genes NDUFB3, NGLY1 and SLC25A4 were highly expressed in RA. It can be used as a potential biomarker for RA.15 It has also been found that artemisinin can enhance the lactylation modification of pyruvate kinase M2 (PKM2) in FLSs. Moreover, this effect further promotes the binding of modifying enzyme of lactylation p300, induces cell cycle arrest in the S phase, regulates the cell cycle, and ultimately inhibits RA-mediated synovial hyperplasia and inflammation.16 In view of the critical mechanism of LA and lactylation, in this paper, we review whether lactylation can be a potential therapeutic target for RA through the research on lactylation in RA-related cells and mechanisms and provide novel perspectives on RA development, identification, and management.
PTMs and RA
PTM refers to adding biochemical components to most proteins during or after protein translation. This process is crucial for regulating protein structure, localisation, and function and is characterised by its ability to change and be reversed. PTMs add a unique level of complexity to the proteome that is not dependent on DNA. They act as continuously fine-tuned regulators in various cellular processes.17 PTMs such as citrullination, carbamylation, acetylation, ubiquitination, and methylation have been associated with RA development.18
As is well-known, citrullinated proteins are critical in the pathogenesis of RA. Studies have found that citrulline-specific helper T cell 1 (TH1) and TH17 cells are increased in patients with RA.19,20 The synovial biopsy samples from people with RA have shown a considerably more significant amount of citrullinated protein compared to the synovium of healthy individuals.21 Anti-carbamylated proteins (anti-CarP) are specific indicators for the diagnosis of RA, and their production is related to the carbamylation of Homocitrulline.22,23 Autophagy was also found to correlate with carbamylation levels in mononuclear cells from treatment-naïve patients with RA.24
In addition, it has been found that the dysregulation of acetylation can hinder the progression of RA by decreasing the expression of forkhead box protein P3 (FOXP3),25 and histone acetylation shows cross-reaction with ACPA in seronegative patients with RA.26 In addition, ubiquitination can also affect the signalling of signalling factors such as tumour necrosis factor (TNF) receptor-associated factor-2 (TRAF2), TRAF6, which are directly or indirectly involved in pro-inflammatory activity and inflammation, thus aggravating the symptoms of RA.27 Methylation modifications, including DNA and RNA methylation, are also associated with RA development. Global DNA hypomethylation has been observed in peripheral blood mononuclear cells (PBMCs) and FLS of individuals with RA.28 The hypomethylated sites are detected in crucial genes associated with RA, such as signal transducer and activator of transcription 3 (STAT3).29 RNA methylation has been implicated in the pathogenesis of RA in macrophages and FLS. It was found that activation of lipopolysaccharide (LPS) can increase the expression of methyltransferase-like 3 (METTL3) in macrophages and increase the total amount of N6-methyladenosine (m6A).30 METTL3 can enhance the inflammatory response of RA FLS by stimulating the nuclear factor-κB (NF-κB) signaling pathway.31
Drugs for PTMs, such as proteasomal inhibitors, are also being developed and tested in clinical trials. The proteasomal inhibitor has the ability to exert its effects at several levels, suppressing transforming growth factor-β (TGF-β)-activated kinase 1 (TAK1) and TRAF6 signalling pathways, possibly rendering cancerous cells prone to being affected by chemotherapy and programmed cell death. They are also thought to be anti-inflammatory agents, having, under normal conditions, been shown to block IL, cell-sticky molecules, and enzymes.32–34 Bortezomib is a proteasomal inhibitor extensively studied in pre-clinical trials. It effectively inhibits the 26S proteasome, resulting in reduced cell proliferation and the induction of chemotherapy. Bortezomib is commonly used in the treatment of malignancies and arthritis. It is authorised by the Food and Drug Administration (FDA), effectively hinders IĸB kinase and aids in preventing the breakdown of NF-κB. This action is crucial in managing inflammatory illnesses.35–37 Moreover, Bortezomib has been reported that it can improve joint performance for RA.38
Evidently, many types of PTM have been found to have specific functions in the onset and progression of RA, so does the newly emerged PTM lactylation mediated by the metabolite LA also play a particular role in RA? Continuing to explore this will help to clarify the aetiology of RA and the investigation of novel pharmaceuticals, and it is believed that the exploration of the related mechanism in the lactylation will also bring new enlightenment for RA.
LA and RA
LA mainly exists in the body as two enantiomers, namely L-LA and D-LA. Although L-LA is a frequently occurring byproduct of the body’s metabolic processes, D-LA is only created in small amounts by some microbes.39 In humans, L-LA primarily arises from converting glucose and alanine into pyruvate.40 Glucose undergoes glycolysis and the pentose phosphate pathway to produce pyruvate. Lactate dehydrogenase (LDH) catalyses the reduction of pyruvate to L-LA.41 LA serves as both a fuel for cells and perhaps contributes to the development of certain diseases, as well as the regulation of cell activity.42,43 The potential of LA as a signalling molecule was largely overlooked until 2008, when a groundbreaking report emerged to enhance our understanding of LA-mediated signal transduction. Ge et al were the pioneers in discovering GPR81, a G protein-coupled receptor (GPCR) that interacts explicitly with LA and transmits signals into cells.44 When GPR81 is combined with LA, α-subunits with Guanosine triphosphate-Guanosine diphosphate (GTP-GDP) immediately switch energy structure transformation occurs, which will affect the downstream signalling molecules, such as cyclic adenosine monophosphate (cAMP) and Ca2+.45
Hypoxia is a fundamental metabolic change in many inflammatory diseases, and the anaerobic glycolysis pathway must provide an adenosine triphosphate (ATP) supply to sustain the energy needed for rapid cell growth. This statement aligns with previous research investigations on RA that have reported increased amounts of LA and reduced glucose in the synovial fluid.46–48 According to reports, heightened glycolytic activity and elevated levels of LA and pyruvate contribute to the development of angiogenesis and pannus production.49 These processes ultimately lead to the destruction of joint tissue structure in RA. LA also enhances activation and movement while promoting the growth of proinflammatory cytokines like TNF and IL, which are released by immune cells. TNF and IL-1 are the main inflammatory factors involved in RA. LA enhances the process of activating the transcription of TNF and IL-1 by increasing the transcriptional activity of NF-κB. Furthermore, LA plays a crucial function in relation to myeloid differentiation factor-2 (MD-2), the stimulation of the Toll-like receptor 4 (TLR4) community, and the activation of inflammatory genes in human U937 tissue cells.50 Moreover, LA can be especially involved in the onset and evolution of RA by controlling the differentiation of T cells, macrophages, B cells, mesonuclear cells, and other cells and the secretion of inflammatory factors.51 At present, many monocarboxylate transporters (MCT) inhibitors, including bindarit,52 phloretin,53 and other drugs regulating LDH activity such as FX11,54 GSK2837808A55 have been developed to treat tumour and arthritis by regulating LA. Research has demonstrated that these medications play a beneficial function in the management and enhancement of RA by regulating the transit and buildup of LA.
Lactylation and the Effect of Lactylation on Diseases
In addition to the direct role of LA in the pathogenesis of the disease, it has been found that PTM lactylation mediated by LA may play a role in regulating gene expression (the process of lactylation is shown in Figure 1). Building upon the identification of different histone acylations originating from cellular metabolites, Zhang and his team hypothesised and confirmed the existence of lysine lactylation as a novel kind of histone modification that can be induced by LA.10 The study utilised mass spectrometry to identify all four canonical histones that undergo lysine lactylation in bone marrow-derived macrophages (BMDMs) and HeLa cells. Subsequently, Wan et al employed the cyclic immonium (cycIm) ion to detect previously unknown lactylated proteins and modification sites accurately. Additionally, they uncovered a more comprehensive range of lactylation occurrences in the human proteome, extending beyond histones. They took advantage of multiple publicly accessible proteome datasets and made interesting observations. They noted that the identified lactylation proteins were not confined to the nucleus but were also found in the cytoplasm. Among them, many enzymes involved in glycolysis, such as fructose bisphosphate aldolase A (ALDOA), were heavily lactylated.56
With the discovery of lactylation, more and more studies have confirmed that it is indeed involved in developing many diseases. In terms of tumours, the study demonstrates that there is an increase in histone lactylation in tumours, which is linked to an unfavourable prognosis for ocular melanoma. Histone lactylation mechanistically contributes to the development of tumours by promoting the production of YTH N6-methyladenosine RNA binding protein F2 (YTHDF2). YTHDF2 recognises the m6A-modified period circadian regulator 1 (PER1) and tumour protein P53 (TP53) mRNAs and promotes their degradation, which accelerates tumorigenesis of ocular melanoma.57 Concerning the nervous system, studies suggest that changes in LA levels align with lactylation in brain cells, which is regulated by neuronal excitation and social stress. In the stress model, these stimuli raise lactylation, which is linked with the expression of the neuronal activity marker c-Fos, as well as with reduced social behaviour and increased anxiety-like behaviour. Furthermore included in the study are 63 potential lysine-lactylated proteins and evidence showing that stress preferably enhances histone H1 lactylation.58 Also, a study showed that both 5XFAD mice and individuals with Alzheimer’s disease (AD) detected elevated histone lactylation in brain samples, and the levels of H4K12 lactylation are elevated in microglia adjacent to Aβ plaques. The histone modification dependent on LA is highly concentrated at the promoters of genes involved in glycolysis and enhances transcription, leading to an increase in glycolytic activity.59 Additionally, in polymicrobial sepsis, the study found that macrophages can uptake extracellular LA via MCTs to promote high mobility group box 1 (HMGB1) lactylation via a p300/cyclic adenosine monophosphate response element binding protein (CBP)-dependent mechanism.60
Consequently, Zhou et al deepened their comprehension of the link between inflammation and histone lactylation.61 Encouragingly, the latest single-cell sequencing results found that RA lactylation score was significantly increased in patients with RA, and the study also confirmed that core lactylation-promoting genes NDUFB3, NGLY1, and SLC25A4 were highly expressed in RA, indicating that they may be potential biomarkers of RA.15 Given the above key roles of PTM and LA in RA, and the new findings of lactylation in RA, LA-mediated lactylation of PTM may indeed play a key role in RA. We hope to explore the possible relationship between lactylation and RA from the relevant cells and mechanisms and provide clues and ideas for future mechanism research (the network of relationships among PTMs, lactylation, LA, and RA is shown in Figure 2).
RA-Associated Cells and Lactylation
Lactylation in Macrophages
Macrophages are multifunctional cells that protect the body from infection, injury, autoimmune diseases, or cancer by removing invading pathogens, cancer, and senescent cells and by remodelling or repairing damaged tissues.62 Activated macrophages are classified into pro-inflammatory M1 and anti-inflammatory M2 macrophages according to their function.63 The imbalance of the M1/M2 type, especially the dominance of M1 macrophages, is considered an important pathogenic mechanism of RA. Studies64 have shown that M1 macrophages accumulate in the synovial tissue of patients with RA, constantly play a pro-inflammatory role, damage synovial tissue, and lead to cartilage and bone damage. In peripheral blood, macrophage M1 type is also shown to increase continuously, leading to increased inflammation and aggravation of disease.65 From the perspective of metabolism, M1 macrophages preferentially use glycolysis, while M2 macrophages rely on oxidative phosphorylation.66 The formation of synovium “pus” and the existence of an anoxic inflammatory environment greatly increase the glycolytic activity of macrophages, and macrophages are polarised toward the M1 phenotype, promoting large-scale oxidative tissue damage.67 Therefore, regulating the functional state of macrophages can effectively improve the inflammatory microenvironment, promote tissue repair and restore normal function, and delay the progression of RA disease.
In macrophages, an essential cell of RA,68 the epigenetic regulation of lactylation on macrophages may largely influence RA’s evolution. Studies10 have found that histone lactylation can regulate the function of macrophages through gene expression regulation. In the late stage of polarisation of M1 macrophages, histone lactylation level is significantly increased on the M2 gene promoter and directly activates chromatin gene transcription, promoting the expression of repair genes involved in wound healing (such as Arg1). It was proposed that the transfer of pro-inflammatory M1 to anti-inflammatory M2 macrophages was greatly aided by histone lactylation. Furthermore, the increase of LA leads to the enhancement of histone lactylation modification in macrophages, thus inhibiting the formation of nucleotide-binding oligomerisation domain-like receptor protein 3 (NLRP3) inflammasome in macrophages and reducing the scorch death of macrophages, hence alleviating the inflammatory response.69 In addition to the above-related mechanisms, histone lactylation can also affect the antigen-presenting and immunosuppressive function of macrophages and the immune regulatory process. In a study70 on the treatment of glioblastoma, it was found that LA accumulation up-regulated the expression of nucleoside tri phosphodiesterase CD39, nucleotidase CD73, chemokine receptor 8 (CCR8) in LA-treated cells and macrophages, and directly improved the activity of these gene promoters through histone H3K18 lactylation. They were blocking immune surveillance at the cellular level. Subsequently, studies71,72 have shown that lactylation modification of macrophages can participate in immune regulation, tissue repair, regeneration, tumour immune regulation, inflammatory diseases, and other processes. Given the crucial role of macrophages in RA, these findings have important implications for RA’s mechanism exploration and treatment strategies.
Lactylation in T Cells
T lymphocytes are derived from bone marrow hematopoietic stem cells and mature in the thymus, so they are also called “thymus-dependent lymphocytes” and are an important driver of inflammation and immune response.73 T cells are classified as CD4+T and CD8+T, where CD4+T comprises helper T cells (THs) and regulatory T cells (Tregs). T lymphocyte-mediated immune response has an important impact on the pathological mechanism of RA, and a large number of T cells can be detected in the inflammatory synovium.74 Studies75,76 have reported that TH cells and their secreted cytokines can cause T cell immune regulation disorders, thus promoting the occurrence of RA. Among them, TH1-secreted IL-277 and TH2 cell-produced IL-4, IL-6, and IL-10 can both mediate inflammatory response.78 TH17 cells can promote the proliferation and activation of periosteal fibroblasts through the production of IL-17 and IL-22, produce chemokines, and promote synovial inflammation. However, the abnormal number and (or) function of Treg cells will cause an immune cascade reaction, resulting in increased expression of IL-2 in vivo, thus activating macrophages in the synovial membrane of bone and joint, producing various inflammatory cytokines such as IL-1, IL-6, and IL-8, destroying articular cartilage and aggravating RA.79 Therefore, intervention of T cells has become a meaningful way to treat RA.80
In recent years, the role of glycolytic product LA-mediated PTM lactylation in T cells has gradually attracted attention. Specific metabolic differences play an important role in altering the fate of CD4+T cell differentiation and maintaining appropriate immune function. It was found that the expression of glycolytic metabolism was increased in Th1, Th2, and Th17 cells differentiated in vitro, while glycolysis could limit the immunosuppressive function of Treg cells.81 Glycolysis and elevated LA concentration are well-known features of the microenvironment of inflammatory tissues.82 The upregulation of LA concentration in the synovium of patients with RA in a state of hypoxic inflammation may induce high lactylation of crucial proteins, thereby promoting the function of effector T cells and increasing the production of pro-inflammatory cytokine IL-17.83 Studies have shown that Th17/Treg plasticity in CD4+T cell phenotype is affected by LA, and extracellular LA treatment promotes the development of Treg by enrichment of H3K18 lactylation at the Foxp3 site of Th17 cells.84 In the study85 of CD8+T cells, it was found that after the downregulation of H3K18 lactylation and H3K9 lactylation mediated by lactate dehydrogenase A(LDHA) inhibitors, the effector function of CD8+T cells was weakened, indicating that histone lactylation labelling performs a pivotal function in controlling the transcriptional, metabolic and functional spectrum of CD8+T cell subsets. In tumour-related studies,86 it was discovered that LA increased TGF-β signalling in Tregs and controlled the lactylation of Moesin, which in turn promoted carcinogenesis. Unfortunately, most of the current research on lactylation and T cells comes from the field of oncology. Among autoimmune-related diseases, a study87 of autoimmune uveitis (AU) found that metabolic reprogramming from oxidative phosphorylation to glycolysis and LA accumulation was involved in CD4+T cell differentiation and LA-derived lactylation regulated CD4+T cell differentiation, and its lactylation level increased with the progression of AU. It is suggested that regulating lactylation may be a potential therapeutic target for autoimmune diseases. These studies indicate that not only glycolytic metabolism affects the fate of T cells, but also lactylation mediated by metabolite LA can act on T cells, and it is not difficult to speculate that regulating lactylation in T cells can improve RA.
Lactylation in Osteoblasts
Mesenchymal stem cells, from which osteoblasts are produced, are crucial for the development and repair of new bone. Abnormal function of osteoblasts can lead to osteoporosis, delayed fracture healing, brittle bone disease, etc. Studies have shown that about 80% of patients with RA will have osteoporosis or even fracture,88 and bone destruction caused by RA has been shown to be related to the formation and inhibition of osteoblasts.89
Lactylation has an essential effect on osteoblasts. It was found that after histone H3K18 lactylation in bone marrow mesenchymal stem cells, osteoblast genes that can promote osteoblast differentiation were detected,90 providing support for the regulation of osteoblasts by histone lactylation. LDHA expression, intracellular LA, and histone lactylation levels increased gradually during osteogenic differentiation. JunB is an activating protein-1 transcription factor, and LDHA regulates JunB expression through histone lactylation modification. Studies have shown that JunB expression is decreased in cells with LDHA knockdown. From a mechanism point of view, the knockdown of LDHA reduces the enrichment of histone lactylation markers on the JunB promoter. H3K18 lactylation levels were significantly reduced, and osteogenic differentiation was disrupted.91 Further studies have shown92 that LA synthesised by LDHA during glucose metabolism can promote osteoblast differentiation of C2C12 cells, whereas low LA level leads to less osteoblast differentiation. In addition, p300 silencing also inhibits osteoblast differentiation and reduces histone lactylation. Wnt pathway activation and increased lactylation levels are strongly linked to the potential mechanism of lactylation’s influence on osteogenesis. Studies have found that proanthocyanidins may activate the Wnt pathway by increasing the level of lactylation in inflammatory states, thus promoting the osteogenesis of periodontal ligament stem cells (PDLSC).93 In conclusion, LA and its derived PTM have an essential effect on osteoblasts. LA produced during glucose metabolism can regulate the differentiation of undifferentiated cells into osteoblasts through histone modification epigenetics. However, there are few reports on the regulation of lactylation on osteoblast differentiation. The mechanism of the influence of lactylation on osteoblasts in RA bone destruction and secondary osteoporosis needs further exploration.
RA-Associated Mechanisms and Lactylation
Lactylation in Angiogenesis
One of the several pathways contributing to the pathophysiology of RA is pathological angiogenesis. Synovial tissues of patients with RA stimulated by chronic inflammation release various pro-angiogenic factors, which stimulate vascular endothelial cells to proliferate, differentiate, migrate, and promote neoangiogenesis. Neovascularisation promotes leukocyte recruitment and synovial tissue proliferation by increasing vascular permeability and more oxygen and nutrient supply. Subsequently, abnormally proliferating synoviocytes, inflammatory cells, and neovascularisation constitute an aggressive vascular opacification, which adheres to the surface of the articular cartilage, causing irreversible damage and ultimately leading to joint destruction and dysfunction.94
In the early stage of RA, energy metabolism is characterised by aerobic glycolysis,95 and several studies have shown that LA, as the end product of aerobic glycolysis, is closely related to neovascularisation. Tumour-derived LA promotes endothelial cell activation and angiogenesis.96 In the retina, LA upregulates vascular endothelial growth factor expression in macrophages and promotes choroidal neovascularisation.97 Bao et al found that both endogenous and exogenous nitric oxide (NO) induced by immune activation can induce joint angiogenesis in adjuvant-induced arthritis (AIA) rats by driving synovial hypoxia, in which the levels of NO and LA in the serum of arthritis model mice fluctuated proportionally.98
LA produced by the endogenous glycolytic pathway is a crucial regulator of histone lactylation modification, and when LA produced by glycolysis increases, lysine lactylation also increases. It has been shown that HIF-1α lactylation mediated by MCT1 under normoxic conditions promotes the transcriptional activity of KIAA 1199, which enhances angiogenesis in prostate cancer.99 Wuchererine impairs HIF-1α histone lactylation to inhibit Semaphorin 3A-mediated angiogenesis by inducing iron apoptosis in prostate cancer.100 And in a study of the mechanism of retinal vascular neogenesis, the up-regulation of p300 expression led to elevated microglia YY1 lactylation modification, which in turn enhanced the transcription and expression of fibroblast growth factor 2 (FGF2), thereby promoting angiogenesis.101 In another study, LA was found to exacerbate microvascular abnormalities in diabetic retinopathy by promoting lactylation of the histone H3K18 locus to drive the expression of the obesity-associated protein FTO.102 Therefore, it is possible that lactylation is an important mechanism involved in angiogenesis in RA. Inhibition of neovascularisation by hindering lactylation may be effective in curbing the development of synovitis and ameliorating the arthritic symptoms of RA.
Lactylation in Cell Proliferation, Invasion, and Migration
The proliferative pannus’s main cell type, FLS, serves as the joint destruction’s principal effector cell. RA-FLS have tumour-like cell proliferation and prolonged survival time, which can induce synovial proliferation and pannus formation.103 Studies have shown that sustained proliferation of FLS caused by insufficient apoptosis and excessive glycolysis plays a key role in RA pathophysiology and that an abnormal increase in glycolysis leads to localised hypoxia and an acidic microenvironment in RA joints, which promotes elevated levels of LA, thus increasing the proliferation, invasiveness, and migration rate of FLS and exacerbating cartilage and bone destruction.
Similarly, an abnormal fibroblast phenotype and a hypoxic and nutrient-deficient microenvironment in the pannus are features also found in solid tumours. LA has now been shown to be a critical metabolite that promotes tumour progression, and the formation of an immune-suppressive tumour microenvironment, and then lactylation modifications modulate tumour cell proliferation, migration, and invasion in a wide range of cancers, ultimately leading to malignant progression.104 For instance, lactylation in hepatocellular carcinoma cells stimulates the cell lines’ motility and proliferation.105 In non-small cell lung cancer cells, hypoxia can mediate SOX9 lactylation modification by promoting glycolysis, which supports the migration and invasion of non-small cell lung carcinoma cells.106 And the addition of exogenous LA to LDHA/LDHB-deficient cells of renal clear cell carcinoma restores the level of histone lactylation of the cancer cells and activate the expression of platelet-derived growth factor recepterβ (PDGFRβ), thus restoring their proliferation and migration ability.107 Meanwhile, it was found that LA promoted cell proliferation, migration, and invasion by targeting HMGB1 in endometriosis through the induction of H3K18 lactylation.108 In the study of FLS, it was found that ART, a natural bioactive molecule extracted from artemisia annua, could directly inhibit the key regulator of glycolysis pyruvate kinase PKM2 through p300-mediated lactylation and reduce its proliferation function on FLS.16
From this, it can be seen that lactylation has a crucial role in influencing cell proliferation, migration, and invasion. The discovery of the lacylation mechanism in FLS by Wang et al16 further suggests that the change of lacylation modification of RA-FLS cells may indeed participate in the pathogenesis of RA, and targeting the FLS lactylation modification sites for the treatment of RA may be able to prevent the pathologic development of RA more directly.
Lactylation in Fibrosis
Lactylation not only has a profound influence on the pathogenesis associated with RA but also perhaps plays a key role in RA complications. Up to 6.8% of deaths in women and 9.8% of deaths in men with RA are caused by interstitial lung disease (ILD), one of the most prevalent and potentially severe extra-articular features of RA. ILD typically presents as a radiographic and pathological pattern of generalised interstitial pneumonitis, which is similar to IPF in terms of its pathogenesis and disease behaviour. At the same time, pulmonary fibrosis is a typical pathological feature of a large group of ILD that can lead to the destruction of normal lung structure, lung scarring, and eventual organ failure.109
Lung macrophages and lung fibroblasts are important in the local inflammatory response to pulmonary fibrosis. In the fibrotic microenvironment of lung macrophages, TGF-β-induced lung fibroblasts and bronchoalveolar lavage fluid (BALF) from TGF-β- or bleomycin-induced pulmonary fibrosis mice showed a significant increase in LA content and promoted the lung fibrosis genes PDGFA, thrombospondin 1 (THBS1) and vascular endothelial growth factor A (VEGFA) by upregulating p300-mediated macrophage histone lactylation modification.110 Arsenic exposure induced elevated levels of LA in the microenvironment of lung fibrosis tissues, and alveolar epithelial cells further promoted elevated levels of histone H3K18 lactylation through LA uptake, which activated the neuronal regeneration related protein (NREP)/TGF-β1 signalling pathway, stimulated further activation of lung fibroblasts differentiation, and promoted the disease progression of arsenic-induced pulmonary fibrosis.111 Li et al found that PM2.5 exposure significantly increased LDH levels in murine LDH activity and LA content in lung macrophages and increased the expression of pro-fibrotic mediators by triggering glycolysis in macrophages to induce histone lactylation at the macrophage gene promoter as well as the secretion of pro-fibrotic cytokines.112 These findings further illuminate the pathophysiology of lactylation’s crucial role in the aetiology of pulmonary fibrosis. Meanwhile, histone lactylation is vital in promoting hepatic, placental, and cardiac fibrosis. The induction of liver fibrosis is believed to be largely dependent on the activation and phenotypic modification of quiescent hepatic stellate cells (HSC). It was shown that hexokinase 2 expression boosts glycolytic activity and produces a significant amount of LA, a substrate for histone lactylation, which mediates the activation of H3K18 lactylation to change the active phenotype of HSC and encourages the onset of hepatic fibrosis.113 Zhou et al discovered that to increase HSC activation and liver fibrosis, the m6A reader IGF2BP2 modulates histone lactylation and controls glycolytic metabolism.114 In preeclamptic placentas, hypoxia increases the expression of pro-placental fibrosis factors FN 1 and SERPINE 1 through LA-induced histone lactylation.115 Another study found that after myocardial infarction, LA was able to induce lactylation and nuclear translocation of Snail 1, which promoted endothelial-to-mesenchymal transition, increasing cardiac fibrosis through activation of the TGF-β/Smad 2 signalling pathway.116
A national study conducted in Denmark noted that 14% of RA-ILD cases were diagnosed with lung disease 1–5 years before RA diagnosis and elucidated that lung inflammation plays a central role in RA disease pathogenesis.117 Biomarkers for screening RA-ILD are not currently available in clinical practice, and their treatment criteria have not been standardised. The treatment of RA is a double-edged sword. On the one hand, it can alleviate the progression of RA-ILD. Still, at the same time, it also has the drawbacks of inducing pulmonary toxicity and increasing susceptibility to infection. Investigating the possible function of lactylation in various physiological and pathological processes may offer a novel approach for the diagnosis and treatment of RA-ILD, considering the importance of lactylation in fibrosis of the lung and other diseases’ fibrotic pathology (lactylation and critical cells and essential mechanisms of RA are summarized in Figure 3).
Perspective and Conclusion
Although we have yielded new findings about lactylation, we must acknowledge that there is still much to learn about this subject and that much research on lactylation has not been fully explained.
According to Zhang and associates, histone lactylation is a response to levels of LA and is brought on by the accumulation of LA.10 However, it is unknown if histone lactylation results from LA analogues or is an inevitable consequence of LA buildup. Whether LA concentrations in the nucleus cause histone lactylation is also uncertain. Moreover, palmitoylation takes place at threonine residues and succinylation at cysteine residues.118 Whether lactylation happens at residues of amino acids other than lysine is also unknown. At the same time, the lactylation detection of the modification and the low abundance of lactylated peptides are likely to be overshadowed by the undecorated peptide mass in mass spectrometry. However, it is believed that future use of diagnostic cycIm ions in MS/MS spectroscopy, as well as the use of more publicly accessible datasets, will help to identify relevant lactylation events.119
In terms of epigenetic tools, enzymes that read, write, and erase lactylation are not fully revealed. More evidence has confirmed that p300 is a potential writer of lactylation. In p300-knockdown macrophages, both histone lactylation and profibrotic gene expression were reduced.110 In HEK293T cells, a modest increase in lactylation was observed after p300 overexpression.10 In addition to p300, other proteins have been found to be involved in writing. Through the silent p300/CBP, HMGB1 lactylation was weakened.60 In erasing lactylation modification, Zu et al reported sirtuin 2 (SIRT2) as a potent histone lactylation eraser in vitro.120 In addition, histone deacetylase 1–3 (HDAC1-3) and SIRT1-3 were identified as powerful erasers for lactylation in vitro.121 Through overexpression and knockdown analysis, HDAC1 and HDAC3 were proved to have distinct functions in histone lactylation. Subsequently, HDAC2, HDAC3, HDAC8, SIRT2, and SIRT3 were also regarded as potential lactylation erasers.122 At present, no study has revealed a reading of lactylation. Proteomic analysis of H3K18 lactylation immunoprecipitation assay by Hu et al revealed specific brahma-related gene 1 (Brg1) recruitment during reprogramming, with both H3K18 lactylation and Brg1 enriched at the promoter of genes involved in pluripotency and epithelial junction. The binding of Brg1 to H3K18 lactylation was revealed for the first time, suggesting that it may be a reader of histone lactylation.123 Epigenetic tool enzymes are crucial in the process of lactylation. Given that the research on critical enzymes has made much progress in the acetylation-targeted therapy of tumours,124 identifying the critical enzymes of lactylation may also provide new ideas for RA-targeted therapy.
Lactylation modification acts on the upstream and downstream of the mechanism and interacts with other PTMs. Gu et al found that histone lactylation enhances alpha-ketoglutarate-dependent dioxygenase homolog 3 (ALKBH3) expression and simultaneously attenuates the formation of tumour-suppressive promyelocytic leukaemia protein (PML) condensates by removing the m1A methylation of SP100A, promoting the malignant transformation of cancers.125 Moreover, scientists have discovered that increased doses of LA can induce the acetylation of additional lysine sites. Through the suppression of SIRT1, a known “eraser” accountable for acetylation, this process is promoted.126 Later studies on murine BMDMs revealed that crucial phases in the LA-driven process include the phosphorylation and subsequent degradation of YAP and LATS1, upstream molecules of SIRT1.60 It can be seen that lactylation modification is complex in the process of protein modification, and the research on it is more hopeful of bringing more updates and breakthroughs in epigenetic crosstalk.
Through existing studies, it can be found that a variety of cells, including macrophages and a variety of disease-related pathological mechanisms, can be widely affected by lactylation, and these cells and mechanisms are also closely related to the occurrence and development of RA. At the same time, the mediator of lactylation, LA, also plays a promoting role in RA, revealing the possibility of lactylation in RA. It provides new ideas for researchers. Numerous medications that target histone PTMs have been used in clinical settings to treat a range of illnesses and have shown promising results. For example, medicines that increase histone crotonylation, decrease histone deacetylation by inhibiting HDACs and decrease histone methylation by inhibiting histone methyltransferases are useful for renal injury.127 Therefore, studying the specific process of lactylation to find accurate targets is expected to become a promising treatment for RA. Shortly, more research will eventually solve the mysteries of lactylation, usher in updates in the field of RA and even more other diseases, and then be applied to clinical, bringing new hope to more patients with diseases such as RA.
Abbreviations
LA, lactic acid; PTM, posttranslational modification; RA, rheumatoid arthritis; RF, rheumatoid factor; ACPAs, anti-citrullinated protein antibodies; anti-CarP, anti-carbamylated protein; FOXP3, forkhead box protein P3; TNF, tumor necrosis factor; FLS, fibroblast-like synoviocytes; m6A, N6-methyladenosine; METTL3, methyltransferase-like 3; IL-6, interleukin-6; TGF-β, transforming growth factor-β; TAK1, transforming growth factor-β-activated kinase 1; GPCR, G protein-coupled receptor; cAMP, cyclic adenosine monophosphate; MD-2, myeloid differentiation factor-2; TLR4, Toll-like receptor 4; MCT, monocarboxylate transporters; BMDMs, bone marrow-derived macrophages; CycIm, cyclic immonium; ALDOA, aldolase A; YTHDF2, YTH N6-methyladenosine RNA binding protein F2; PER1, period circadian regulator 1; HMGB1, high mobility group box 1; CBP, cyclic adenosine monophosphate response element binding protein; KLF4, Kruppel-like factor 4; NLRP3, nucleotide-binding oligomerization domain-like receptor protein 3; CCR8, chemokine receptor 8; PDLSC, periodontal ligament stem cells; AIA, adjuvant-induced arthritis; HIF-1α, hypoxia-inducible factor-1α; FGF2, fibroblast growth factor 2; PDGFRβ, platelet-derived growth factor recepterβ; ILD, interstitial lung disease; BALF, bronchoalveolar lavage fluid; THBS1, thrombospondin 1; NREP, neuronal regeneration related protein; HSC, hepatic stellate cells; SIRT2, sirtuin 2; VEGFA, vascular endothelial growth factor A; Brg1, brahma-related gene 1; ALKBH3, alpha-ketoglutarate-dependent dioxygenase homolog 3; PML, promyelocytic leukaemia protein.
Data Sharing Statement
No data was used for the research described in the article.
Compliance with Ethics Guidelines
This manuscript does not involve any research protocols requiring approval by any relevant institutional review boards or ethics committees.
Acknowledgments
Thanks for the support of the Beijing Municipal Natural Science Foundation [No. 7242261], Capital’s Funds for Health Improvement and Research (CFH) [No.2024-1-4162] and the National Natural Science Foundation of China (NSFC) [No. 82074408]. Thanks to BioRender.com.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Giannini D, Antonucci M, Petrelli F, Bilia S, Alunno A, Puxeddu I. One year in review 2020: pathogenesis of rheumatoid arthritis. Clin Exp Rheumatol. 2020;38(3):387–397. doi:10.55563/clinexprheumatol/3uj1ng
2. Myasoedova E, Crowson CS, Kremers HM, Therneau TM, Gabriel SE. Is the incidence of rheumatoid arthritis rising? Results from Olmsted County, Minnesota, 1955-2007. Arthritis Rheum. 2010;62(6):1576–1582. doi:10.1002/art.27425
3. Tobón GJ, Youinou P, Saraux A. The environment, geo-epidemiology, and autoimmune disease: rheumatoid arthritis. Autoimmun Rev. 2010;9(5):A288–292. doi:10.1016/j.autrev.2009.11.019
4. Turesson C, O’Fallon WM, Crowson CS, Gabriel SE, Matteson EL. Extra-articular disease manifestations in rheumatoid arthritis: incidence trends and risk factors over 46 years. Ann Rheum Dis. 2003;62(8):722–727. doi:10.1136/ard.62.8.722
5. Xi X, Ye Q, Fan D, et al. Polycyclic aromatic hydrocarbons affect rheumatoid arthritis pathogenesis via aryl hydrocarbon receptor. Front Immunol. 2022;13:797815. doi:10.3389/fimmu.2022.797815
6. Huang J, Fu X, Chen X, Li Z, Huang Y, Liang C. Promising therapeutic targets for treatment of rheumatoid arthritis. Front Immunol. 2021;12:686155. doi:10.3389/fimmu.2021.686155
7. Mittal N, Mittal R, Sharma A, Jose V, Wanchu A, Singh S. Treatment failure with disease-modifying antirheumatic drugs in rheumatoid arthritis patients. Singapore Med J. 2012;53(8):532–536.
8. Klein K, Gay S. Epigenetics in rheumatoid arthritis. Curr Opin Rheumatol. 2012;27(1):76–82. doi:10.1097/BOR.0000000000000128
9. Izzo LT, Wellen KE. Histone lactylation links metabolism and gene regulation. Nature. 2019;574(7779):492–493. doi:10.1038/d41586-019-03122-1
10. Zhang D, Tang Z, Huang H. Metabolic regulation of gene expression by histone lactylation. Nature. 2019;574(7779):575–580. doi:10.1038/s41586-019-1678-1
11. Garcia-Carbonell R, Divakaruni AS, Lodi A, et al. Critical role of glucose metabolism in rheumatoid arthritis fibroblast-like synoviocytes. Arthritis Rheumatol. 2016;68(7):1614–1626. doi:10.1002/art.39608
12. Gobelet C, Gerster JC. Synovial fluid lactate levels in septic and non-septic arthritides. Ann Rheum Dis. 1984;43(5):742–745. doi:10.1136/ard.43.5.742
13. Gan P-R, Wu H, Zhu Y-L, Shu Y, Wei Y. Glycolysis, a driving force of rheumatoid arthritis. Int Immunopharmacol. 2024;132:111913. doi:10.1016/j.intimp.2024.111913
14. Xin Q, Wang H, Li Q, et al. Lactylation: a passing fad or the future of posttranslational modification. Inflammation. 2022;45(4):1419–1429. doi:10.1007/s10753-022-01637-w
15. Fu W, Wang T, Lu Y, Shi T, Yang Q. The role of lactylation in plasma cells and its impact on rheumatoid arthritis pathogenesis: insights from single-cell RNA sequencing and machine learning. Front Immunol. 2024;15:1453587. doi:10.3389/fimmu.2024.1453587
16. Wang YH, Gao P, Wang YQ, Xu LZ, Zeng KW, Tu PF. Small-molecule targeting PKM2 provides a molecular basis of lactylation-dependent fibroblast-like synoviocytes proliferation inhibition against rheumatoid arthritis. Eur J Pharmacol. 2024;972:176551. doi:10.1016/j.ejphar.2024.176551
17. Tarazona OA, Pourquié O. Exploring the influence of cell metabolism on cell fate through protein post-translational modifications. Dev Cell. 2020;54(2):282–292. doi:10.1016/j.devcel.2020.06.035
18. Kwon EJ, Ju JH. Impact of posttranslational modification in pathogenesis of rheumatoid arthritis: focusing on citrullination, carbamylation, and acetylation. Int J Mol Sci. 2021;22(19):10576. doi:10.3390/ijms221910576
19. James EA, Rieck M, Pieper J. Citrulline-specific Th1 cells are increased in rheumatoid arthritis and their frequency is influenced by disease duration and therapy. Arthritis Rheumatol. 2014;66(7):1712–1722. doi:10.1002/art.38637
20. Cianciotti BC, Ruggiero E, Campochiaro C. CD4+ memory stem T cells recognizing citrullinated epitopes are expanded in patients with rheumatoid arthritis and sensitive to tumor necrosis factor blockade. Arthritis Rheumatol. 2020;72(4):565–575. doi:10.1002/art.41157
21. Makrygiannakis D, Revu S, Engström M, et al. Local administration of glucocorticoids decreases synovial citrullination in rheumatoid arthritis. Arthritis Res Ther. 2012;4(1):R20. doi:10.1186/ar3702
22. Bright R, Proudman SM, Rosenstein ED, Bartold PM. Is there a link between carbamylation and citrullination in periodontal disease and rheumatoid arthritis? Med Hypotheses. 2015;84(6):570–576. doi:10.1016/j.mehy.2015.03.006
23. Shi J, van Steenbergen HW, van Nies JA, et al. The specificity of anti-carbamylated protein antibodies for rheumatoid arthritis in a setting of early arthritis. Arthritis Res Ther. 2015;17:339. doi:10.1186/s13075-015-0860-6
24. Manganelli V, Recalchi S, Capozzi A, et al. Autophagy induces protein carbamylation in fibroblast-like synoviocytes from patients with rheumatoid arthritis. Rheumatology. 2018;57(11):2032–2041. doi:10.1093/rheumatology/key174
25. Su Q, Jing J, Li W. Impaired Tip60-mediated Foxp3 acetylation attenuates regulatory T cell development in rheumatoid arthritis. J Autoimmun. 2019;100:27–39. doi:10.1016/j.jaut.2019.02.007
26. Lloyd KA, Wigerblad G, Sahlström P, et al. Differential ACPA binding to nuclear antigens reveals a pad-independent pathway and a distinct subset of acetylation cross-reactive autoantibodies in rheumatoid arthritis. Front Immunol. 2019;9:3033. doi:10.3389/fimmu.2018.03033
27. Dondelinger Y, Darding M, Bertrand MJM, Walczak H. Poly-ubiquitination in TNFR1-mediated necroptosis. Cell Mol Life Sci. 2016;73(11–12):2165–2176. doi:10.1007/s00018-016-2191-4
28. Liu CC, Fang TJ, Ou TT, et al. Global DNA methylation, DNMT1, and MBD2 in patients with rheumatoid arthritis. Immunol Lett. 2011;135(1–2):96–99. doi:10.1016/j.imlet.2010.10.003
29. Hirano T. Interleukin 6 in autoimmune and inflammatory diseases: a personal memoir. Proc Jpn Acad Ser B Phys Biol Sci. 2010;86(7):717–730. doi:10.2183/pjab.86.717
30. Wang J, Yan S, Lu H, Wang S, Xu D. METTL3 attenuates lps-induced inflammatory response in macrophages via NF-κB signaling pathway. Mediators Inflamm. 2019;2019(2019):3120391. doi:10.1155/2019/3120391
31. Shi W, Zheng Y, Luo S, et al. METTL3 promotes activation and inflammation of FLSs through the NF-κB signaling pathway in rheumatoid arthritis. Front Med. 2021;8:607585. doi:10.3389/fmed.2021.607585
32. Tsukamoto S, Yokosawa H. Natural products inhibiting the ubiquitin-proteasome proteolytic pathway, a target for drug development. Curr Med Chem. 2006;13(7):745–754. doi:10.2174/092986706776055571
33. Elliott PJ, Zollner TM, Boehncke WH. Proteasome inhibition: a new anti-inflammatory strategy. J Mol Med. 2003;81(4):235–245. doi:10.1007/s00109-003-0422-2
34. Richardson PG, Mitsiades C, Hideshima T, Anderson KC. Bortezomib: proteasome inhibition as an effective anticancer therapy. Annu Rev Med. 2006;57:33–47. doi:10.1146/annurev.med.57.042905.122625
35. Nawrocki ST, Carew JS, Dunner K Jr, et al. Bortezomib inhibits PKR-like endoplasmic reticulum (ER) kinase and induces apoptosis via ER stress in human pancreatic cancer cells. Cancer Res. 2005;65(24):11510–11519. doi:10.1158/0008-5472.CAN-05-2394
36. Bogyo M, Wang EW. Proteasome inhibitors: complex tools for a complex enzyme. Curr Top Microbiol Immunol. 2002;268:185–208. doi:10.1007/978-3-642-59414-4_8
37. Palombella VJ, Rando OJ, Goldberg AL, Maniatis T. The ubiquitin-proteasome pathway is required for processing the NF-kappa B1 precursor protein and the activation of NF-kappa B. Cell. 1994;78(5):773–785. doi:10.1016/S0092-8674(94)90482-0
38. Lassoued S, Moyano C, Beldjerd M, Pauly P, Lassoued D, Billey T. Bortezomib improved the joint manifestations of rheumatoid arthritis in three patients. Joint Bone Spine. 2019;86(3):381–382. doi:10.1016/j.jbspin.2019.01.019
39. Talasniemi JP, Pennanen S, Savolainen H, Niskanen L, Liesivuori J. Analytical investigation: assay of D-lactate in diabetic plasma and urine. Clin Biochem. 2008;41(13):1099–1103. doi:10.1016/j.clinbiochem.2008.06.011
40. Perriello G, Jorde R, Nurjhan N, et al. Estimation of glucose-alanine-lactate-glutamine cycles in postabsorptive humans: role of skeletal muscle. Am J Physiol. 1995;269(3 Pt 1):E443–450. doi:10.1152/ajpendo.1995.269.3.E443
41. Lemire J, Mailloux RJ, Appanna VD. Mitochondrial lactate dehydrogenase is involved in oxidative-energy metabolism in human astrocytoma cells (CCF-STTG1. PLoS One. 2008;3(2):e1550. doi:10.1371/journal.pone.0001550
42. Bozzo L, Puyal J, Chatton JY. Lactate modulates the activity of primary cortical neurons through a receptor-mediated pathway. PLoS One. 2013;8(8):e71721. doi:10.1371/journal.pone.0071721
43. Latham T, Mackay L, Sproul D, et al. Lactate, a product of glycolytic metabolism, inhibits histone deacetylase activity and promotes changes in gene expression. Nucleic Acids Res. 2012;40(11):4794–4803. doi:10.1093/nar/gks066
44. Ge H, Weiszmann J, Reagan JD. Elucidation of signaling and functional activities of an orphan GPCR, GPR81. J Lipid Res. 2008;49(4):797–803. doi:10.1194/jlr.M700513-JLR200
45. Husted AS, Trauelsen M, Rudenko O, Hjorth SA, Schwartz TW. GPCR-mediated signaling of metabolite. Cell Metab. 2017;25(4):777–796. doi:10.1016/j.cmet.2017.03.008
46. Naughton D, Whelan M, Smith EC, Williams R, Blake DR, Grootveld M. An investigation of the abnormal metabolic status of synovial fluid from patients with rheumatoid arthritis by high field proton nuclear magnetic resonance spectroscopy. FEBS Lett. 1993;317(1–2):135–138. doi:10.1016/0014-5793(93)81508-W
47. Hitchon CA, El-Gabalawy HS, Bezabeh T. Characterization of synovial tissue from arthritis patients: a proton magnetic resonance spectroscopic investigation. Rheumatol Int. 2009;29(10):1205–1211. doi:10.1007/s00296-009-0865-z
48. Ciurtin C, Cojocaru VM, Miron IM. Correlation between different components of synovial fluid and pathogenesis of rheumatic diseases. Rom J Intern Med. 2006;44(2):171–181.
49. Yang XY, Zheng KD, Lin K, et al. Energy metabolism disorder as a contributing factor of rheumatoid arthritis: a comparative proteomic and metabolomic study. PLoS One. 2015;10(7):e0132695. doi:10.1371/journal.pone.0132695
50. Samuvel DJ, Sundararaj KP, Nareika A, Lopes-Virella MF, Huang Y. Lactate boosts TLR4 signaling and NF-kappaB pathway-mediated gene transcription in macrophages via monocarboxylate transporters and MD-2 up-regulation. J Immunol. 2009;182(4):2476–2484. doi:10.4049/jimmunol.0802059
51. Wang Q, Asenso J, Xiao N, et al. Lactic acid regulation: a potential therapeutic option in rheumatoid arthritis. J Immunol Res. 2022;2022:2280973. doi:10.1155/2022/2280973
52. Futagi Y, Kobayashi M, Narumi K, Furugen A, Iseki K. Identification of a selective inhibitor of human monocarboxylate transporter 4. Biochem Biophys Res Commun. 2018;495(1):427–432. doi:10.1016/j.bbrc.2017.10.025
53. Jones RS, Parker MD, Morris ME, Quercetin M. Luteolin, and phloretin are dietary flavonoid inhibitors of monocarboxylate transporter 6. Mol Pharm. 2017;14(9):2930–2936. doi:10.1021/acs.molpharmaceut.7b00264
54. Souto‐Carneiro MM, Klika KD, Abreu MT. Effect of increased lactate dehydrogenase a activity and aerobic glycolysis on the proinflammatory profile of autoimmune CD8+ T cells in rheumatoid arthritis. Arthritis Rheumatol. 2020;72(12):2050–2064. doi:10.1002/art.41420
55. Li HM, Guo HL, Xu C. Inhibition of glycolysis by targeting lactate dehydrogenase A facilitates hyaluronan synthase 2 synthesis in synovial fibroblasts of temporomandibular joint osteoarthritis. Bone. 2020;141:115584. doi:10.1016/j.bone.2020.115584
56. Wan N, Wang N, Yu S. Cyclic immonium ion of lactyllysine reveals widespread lactylation in the human proteome. Nat Methods. 2022;19(7):854–864. doi:10.1038/s41592-022-01523-1
57. Yu J, Chai P, Xie M, et al. Histone lactylation drives oncogenesis by facilitating m6A reader protein YTHDF2 expression in ocular melanoma. Genome Biol. 2021;22(1):85. doi:10.1186/s13059-021-02308-z
58. Hagihara H, Shoji H, Otabi H. Protein lactylation induced by neural excitation. Cell Reports. 2021;37(2):109820. doi:10.1016/j.celrep.2021.109820
59. Pan RY, He L, Zhang J, et al. Positive feedback regulation of microglial glucose metabolism by histone H4 lysine 12 lactylation in Alzheimer’s disease. Cell Metab. 2022;34(4):634–648.e6. doi:10.1016/j.cmet.2022.02.013
60. Yang K, Fan M, Wang X, et al. Lactate promotes macrophage HMGB1 lactylation, acetylation, and exosomal release in polymicrobial sepsis. Cell Death Differ. 2022;29(1):133–146. doi:10.1038/s41418-021-00841-9
61. Zhou Y, Yang L, Liu X, Wang H. Lactylation may be a novel posttranslational modification in inflammation in neonatal hypoxic-ischemic encephalopathy. Front Pharmacol. 2022;13:926802. doi:10.3389/fphar.2022.926802
62. Wang L, Zhang S, Wu H, Rong X, Guo J. M2b macrophage polarization and its roles in diseases. J Leukoc Biol. 2019;106(2):345–358. doi:10.1002/JLB.3RU1018-378RR
63. Kadomoto S, Izumi K, Mizokami A. Macrophage polarity and disease control. Int J Mol Sci. 2021;23(1):144. doi:10.3390/ijms23010144
64. Culemann S, Grüneboom A, Nicolás-ávila JÁ, et al. Locally renewing resident synovial macrophages provide a protective barrier for the joint. Nature. 2019;572(7771):670–675. doi:10.1038/s41586-019-1471-1
65. Vandooren B, Noordenbos T, Ambarus C, et al. Absence of a classically activated macrophage cytokine signature in peripheral spondylarthritis, including psoriatic arthritis. Arthritis Rheum. 2009;60(4):966–975. doi:10.1002/art.24406
66. O’Neill LA. A broken Krebs cycle in macrophages. Immunity. 2015;42(3):393–394. doi:10.1016/j.immuni.2015.02.017
67. Salisbury AK, Duke O, Poulter LW. Macrophage-like cells of the pannus area in rheumatoid arthritic joints. Scand J Rheumatol. 1987;16(4):263–272. doi:10.3109/03009748709102927
68. Sharma A, Goel A. Pathogenesis of rheumatoid arthritis and its treatment with anti-inflammatory natural products. Mol Biol Rep. 2023;50(5):4687–4706. doi:10.1007/s11033-023-08406-4
69. Sun S, Xu X, Liang L. Lactic acid-producing probiotic saccharomyces cerevisiae attenuates ulcerative colitis via suppressing macrophage pyroptosis and modulating gut microbiota. Front Immunol. 2021;12:777665. doi:10.3389/fimmu.2021.777665
70. Li Y, Liu B, Cao Y, et al. Metformin-induced reduction of CCR8 enhances the anti-tumor immune response of PD-1 immunotherapy in glioblastoma. Eur J Pharmacol. 2024;964:176274. doi:10.1016/j.ejphar.2023.176274
71. Irizarry-Caro RA, McDaniel MM, Overcast GR, Jain VG, Troutman TD, Pasare C. TLR signaling adapter BCAP regulates inflammatory to reparatory macrophage transition by promoting histone lactylation. Proc Natl Acad Sci U S A. 2020;117(48):30628–30638. doi:10.1073/pnas.2009778117
72. Wang N, Wang W, Wang X, et al. Histone lactylation boosts reparative gene activation post-myocardial infarction. Circ Res. 2022;131(11):893–908. doi:10.1161/CIRCRESAHA.122.320488
73. Almutairi KB, Nossent JC, Preen DB, Keen HI, Inderjeeth CA. The prevalence of rheumatoid arthritis: a systematic review of population-based studies. J Rheumatol. 2021;48(5):669–676. doi:10.3899/jrheum.200367
74. Panayi GS. Even though T-cell-directed trials have been of limited success, is there reason for optimism. Nat Clin Pract Rheumatol. 2006;2(2):58–59. doi:10.1038/ncprheum0094
75. Maddur MS, Miossec P, Kaveri SV, Bayry J. Th17 cells: biology, pathogenesis of autoimmune and inflammatory diseases, and therapeutic strategies. Am J Pathol. 2012;181:8–18. doi:10.1016/j.ajpath.2012.03.044
76. Sharma A, Goel A. Inflammatory cytokines in rheumatoid arthritis: diagnostic challenges, pathogenic mechanisms and their role in depression and management. Curr Top Med Chem. 2023;23(27):2535–2551. doi:10.2174/1568026623666230915095151
77. Li S, Wang H, Wu H, Chang X. Therapeutic effect of exogenous regulatory T cells on collagen-induced arthritis and rheumatoid arthritis. Cell Transplant. 2020;29:2138946838. doi:10.1177/0963689720954134
78. Wang Z, Zhuo F, Chu P, Yang X, Zhao G. Germacrone alleviates collagen-induced arthritis via regulating Th1/Th2 balance and NF-κB activation. Biophys Res Commun. 2019;518(3):560–564. doi:10.1016/j.bbrc.2019.08.084
79. Jiang Q, Yang G, Liu Q, Wang S, Cui D. Function and role of regulatory t cells in rheumatoid arthritis. Front Immunol. 2021;12:626193. doi:10.3389/fimmu.2021.626193
80. Mellado M, Martínez-Muñoz L, Cascio G, Lucas P, Pablos JL, Rodríguez-Frade JM. T cell migration in rheumatoid arthritis. Front Immunol. 2015;6:384. doi:10.3389/fimmu.2015.00384
81. Yang J, Chen Y, Li X, et al. Complex interplay between metabolism and CD4+ T-cell activation, differentiation, and function: a novel perspective for atherosclerosis immunotherapy. Cardiovasc Drugs Ther. 2024;38(5):1033–1046. doi:10.1007/s10557-023-07466-9
82. Xu K, Yin N, Peng M, et al. Glycolytic ATP fuels phosphoinositide 3-kinase signaling to support effector T helper 17 cell responses. Immunity. 2021;54(5):976–987.e7. doi:10.1016/j.immuni.2021.04.008
83. Haas R, Smith J, Rocher-Ros V, et al. Lactate regulates metabolic and pro-inflammatory circuits in control of T cell migration and effector functions. PLoS Biol. 2015;13(7):e1002202. doi:10.1371/journal.pbio.1002202
84. Lopez Krol A, Nehring HP, Krause FF, et al. Lactate induces metabolic and epigenetic reprogramming of pro-inflammatory Th17 cells. EMBO Rep. 2022;23(12):e54685. doi:10.15252/embr.202254685
85. Raychaudhuri D, Singh P, Hennessey M, et al. Histone lactylation drives CD8 T cell metabolism and function. Preprint bioRxiv. 2024;2024:554830.
86. Gu J, Zhou J, Chen Q, et al. Tumor metabolite lactate promotes tumorigenesis by modulating MOESIN lactylation and enhancing TGF-β signaling in regulatory T cells. Cell Rep. 2022;40(3):111122. doi:10.1016/j.celrep.2022.111122
87. Fan W, Wang X, Zeng S, et al. Global lactylome reveals lactylation-dependent mechanisms underlying TH17 differentiation in experimental autoimmune uveitis. Sci Adv. 2023;9(42):eadh4655. doi:10.1126/sciadv.adh4655
88. Chung SJ, Kwon YJ, Park MC, Park YB, Lee SK. The correlation between increased serum concentrations of interleukin-6 family cytokines and disease activity in rheumatoid arthritis patients. Yonsei Med J. 2011;52(1):113–120. doi:10.3349/ymj.2011.52.1.113
89. Tsukasaki M, Takayanagi H. Osteoimmunology: evolving concepts in bone-immune interactions in health and disease. Nat Rev Immunol. 2019;19(10):626–642. doi:10.1038/s41577-019-0178-8
90. Wu J, Hu M, Jiang H, et al. Endothelial cell-derived lactate triggers bone mesenchymal stem cell histone lactylation to attenuate osteoporosis. Adv Sci. 2023;10(31):e2301300. doi:10.1002/advs.202301300
91. Nian F, Qian Y, Xu F, Yang M, Wang H, Zhang Z. LDHA promotes osteoblast differentiation through histone lactylation. Biochem Biophys Res Commun. 2022;615:31–35. doi:10.1016/j.bbrc.2022.05.028
92. Minami E, Sasa K, Yamada A, et al. Lactate-induced histone lactylation by p300 promotes osteoblast differentiation. PLoS One. 2023;18(12):e0293676. doi:10.1371/journal.pone.0293676
93. Wu Y, Wang X, Zhang Y, et al. Proanthocyanidins Ameliorate LPS-Inhibited Osteogenesis of PDLSCs by Restoring Lysine Lactylation. Int J Mol Sci. 2024;25(5):2947. doi:10.3390/ijms25052947
94. Marrelli A, Cipriani P, Liakouli V, et al. Angiogenesis in rheumatoid arthritis: a disease specific process or a common response to chronic inflammation. Autoimmun Rev. 2011;10(10):595–598. doi:10.1016/j.autrev.2011.04.020
95. Xu L, Chang C, Jiang P, et al. Metabolomics in rheumatoid arthritis: advances and review. Front Immunol. 2022;13:961708. doi:10.3389/fimmu.2022.961708
96. Colegio OR, Chu NQ, Szabo AL, et al. Functional polarization of tumour-associated macrophages by tumour-derived lactic acid. Nature. 2014;513(7519):559–563. doi:10.1038/nature13490
97. Song J, Lee K, Park SW, et al. Lactic acid upregulates VEGF expression in macrophages and facilitates choroidal neovascularization. Invest Ophthalmol Vis Sci. 2018;59(8):3747–3754. doi:10.1167/iovs.18-23892
98. Bao F, Wu P, Xiao N, Qiu F, Zeng QP. Nitric oxide-driven hypoxia initiates synovial angiogenesis, hyperplasia and inflammatory lesions in mice. PLoS One. 2012;7(3):e34494. doi:10.1371/journal.pone.0034494
99. Luo Y, Yang Z, Yu Y, Zhang P. HIF1alpha lactylation enhances KIAA1199 transcription to promote angiogenesis and vasculogenic mimicry in prostate cancer. Int J Biol Macromol. 2022;222(Pt B):2225–2243. doi:10.1016/j.ijbiomac.2022.10.014
100. Yu Y, Huang X, Liang C, Zhang P. Evodiamine impairs HIF1A histone lactylation to inhibit Sema3A-mediated angiogenesis and PD-L1 by inducing ferroptosis in prostate cancer. Eur J Pharmacol. 2023;957:176007. doi:10.1016/j.ejphar.2023.176007
101. Wang X, Fan W, Li N, et al. YY1 lactylation in microglia promotes angiogenesis through transcription activation-mediated upregulation of FGF2. Genome Biol. 2023;24(1):87. doi:10.1186/s13059-023-02931-y
102. Chen X, Wang Y, Wang J-N. Lactylation-driven FTO targets CDK2 to aggravate microvascular anomalies in diabetic retinopathy. EMBO Mol Med. 2024;16(2):294–318. doi:10.1038/s44321-024-00025-1
103. de Oliveira PG, Farinon M, Sanchez-Lopez E, Miyamoto S, Guma M. Fibroblast-Like synoviocytes glucose metabolism as a therapeutic target in rheumatoid arthritis. Front Immunol. 2019;10:1743. doi:10.3389/fimmu.2019.01743
104. Xie Y, Hu H, Liu M, et al. The role and mechanism of histone lactylation in health and diseases. Front Genet. 2022;13:949252. doi:10.3389/fgene.2022.949252
105. Cai D, Yuan X, Cai DQ, et al. Integrative analysis of lactylation-related genes and establishment of a novel prognostic signature for hepatocellular carcinoma. J Cancer Res Clin Oncol. 2023;149(13):11517–11530. doi:10.1007/s00432-023-04947-0
106. Yan F, Teng Y, Li X, et al. Hypoxia promotes non-small cell lung cancer cell stemness, migration, and invasion via promoting glycolysis by lactylation of SOX9. Cancer Biol Ther. 2024;25(1):2304161. doi:10.1080/15384047.2024.2304161
107. Yang J, Luo L, Zhao C. A positive feedback loop between inactive VHL-Triggered histone lactylation and PDGFRbeta signaling drives clear cell renal cell carcinoma progression. Int J Biol Sci. 2022;18(8):3470–3483. doi:10.7150/ijbs.73398
108. Chen J, Qin P, Sun Y, Han S. Histone lactylation promotes cell proliferation, migration and invasion through targeting HMGB1 in endometriosis. J Biomed Res. 2023;37(6):470–478. doi:10.7555/JBR.37.20230095
109. Diesler R, Cottin V. Pulmonary fibrosis associated with rheumatoid arthritis: from pathophysiology to treatment strategies. Expert Rev Respir Med. 2022;16(5):541–553. doi:10.1080/17476348.2022.2089116
110. Cui H, Xie N, Banerjee S, et al. Lung myofibroblasts promote macrophage profibrotic activity through lactate-induced histone lactylation. Am J Respir Cell Mol Biol. 2021;64(1):115–125. doi:10.1165/rcmb.2020-0360OC
111. Wang P, Xie D, Xiao T, et al. H3K18 lactylation promotes the progression of arsenite-related idiopathic pulmonary fibrosis via YTHDF1/m6A/NREP. J Hazard Mater. 2024;461:132582. doi:10.1016/j.jhazmat.2023.132582
112. Li J, Zeng G, Zhang Z, et al. Urban airborne PM(2.5) induces pulmonary fibrosis through triggering glycolysis and subsequent modification of histone lactylation in macrophages. Ecotoxicol Environ Saf. 2024;273:116162. doi:10.1016/j.ecoenv.2024.116162
113. Rho H, Terry AR, Chronis C, Hay N. Hexokinase 2-mediated gene expression via histone lactylation is required for hepatic stellate cell activation and liver fibrosis. Cell Metab. 2023;35(8):1406–1423. doi:10.1016/j.cmet.2023.06.013
114. Zhou Y, Yan J, Huang H, et al. The m(6)A reader IGF2BP2 regulates glycolytic metabolism and mediates histone lactylation to enhance hepatic stellate cell activation and liver fibrosis. Cell Death Dis. 2024;15(3):189. doi:10.1038/s41419-024-06509-9
115. Li X, Yang N, Wu Y, et al. Hypoxia regulates fibrosis-related genes via histone lactylation in the placentas of patients with preeclampsia. J Hypertens. 2022;40(6):1189–1198. doi:10.1097/HJH.0000000000003129
116. Fan M, Yang K, Wang X, et al. Lactate promotes endothelial-to-mesenchymal transition via Snail1 lactylation after myocardial infarction. Sci Adv. 2023;9(5):c9465. doi:10.1126/sciadv.adc9465
117. Kadura S, Raghu G. Rheumatoid arthritis-interstitial lung disease: manifestations and current concepts in pathogenesis and management. Eur Respir Rev. 2021;30(160):210011. doi:10.1183/16000617.0011-2021
118. Diskin C, Ryan TAJ, O’Neill LAJ. Modification of proteins by metabolites in immunity. Immunity. 2021;54(1):19–31. doi:10.1016/j.immuni.2020.09.014
119. Wu X, Tao WA. Uncovering ubiquitous protein lactylation. Nat Methods. 2022;19(7):793–794. doi:10.1038/s41592-022-01536-w
120. Zu H, Li C, Dai C. SIRT2 functions as a histone delactylase and inhibits the proliferation and migration of neuroblastoma cells. Cell Discov. 2022;8(1):54. doi:10.1038/s41421-022-00398-y
121. Moreno-Yruela C, Zhang D, Wei W, et al. Class I histone deacetylases (HDAC1-3) are histone lysine delactylases. Sci Adv. 2022;8(3):eabi6696. doi:10.1126/sciadv.abi6696
122. Zessin M, Meleshin M, Praetorius L, Sippl W, Bařinka C, Schutkowski M. Uncovering robust delactoylase and depyruvoylase activities of HDAC isoforms. ACS Chem Biol. 2022;17(6):1364–1375. doi:10.1021/acschembio.1c00863
123. Hu X, Huang X, Yang Y, et al. Dux activates metabolism-lactylation-MET network during early iPSC reprogramming with Brg1 as the histone lactylation reader. Nucleic Acids Res. 2024;52(10):5529–5548. doi:10.1093/nar/gkae183
124. Ren C, Wu Q, Xiao R, et al. Expanding the scope of genetically encoded lysine post-translational modifications with lactylation, β-hydroxybutyrylation and lipoylation. Chembiochem. 2022;23(18):e202200302. doi:10.1002/cbic.202200302
125. Gu X, Zhuang A, Yu J, et al. Histone lactylation-boosted ALKBH3 potentiates tumor progression and diminished promyelocytic leukemia protein nuclear condensates by m1A demethylation of SP100A. Nucleic Acids Res. 2024;52(5):2273–2289. doi:10.1093/nar/gkad1193
126. Chelladurai P, Boucherat O, Stenmark K. Targeting histone acetylation in pulmonary hypertension and right ventricular hypertrophy. Br J Pharmacol. 2021;178(1):54–71. doi:10.1111/bph.14932
127. Fontecha-Barriuso M, Martin-Sanchez D, Ruiz-Andres O. Targeting epigenetic DNA and histone modifications to treat kidney disease. Nephrol Dial Transplant. 2018;33(11):1875–1886. doi:10.1093/ndt/gfy009
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The Role and Mechanism of Protein Post‑Translational Modification in Rheumatoid Arthritis
Wen J, Liu J, Wan L, Wang F
Journal of Inflammation Research 2025, 18:9055-9078
Published Date: 11 July 2025