")
Back to Journals » Journal of Inflammation Research » Volume 17
Protective Effects and Mechanisms of Inhibiting Endoplasmic Reticulum Stress on Cold Seawater Immersion Combined with Hemorrhagic Shock
Authors Zhou X, Zou L, Deng H, Zhou Y, Wu Y, Ouyang X, Liu L, Wang L, Li T
Received 7 May 2024
Accepted for publication 11 July 2024
Published 23 July 2024 Volume 2024:17 Pages 4923—4940
DOI https://doi.org/10.2147/JIR.S469622
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Xiaowei Zhou, Liyong Zou, Haoyue Deng, Yuanqun Zhou, Yue Wu, Xingnan Ouyang, Liangming Liu, Li Wang,* Tao Li*
Department of Shock and Transfusion, Army Medical Center of Army Medical University, Chongqing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Li Wang; Tao Li, Department of Shock and Transfusion, Army Medical Center of Army Medical University, Chongqing, People’s Republic of China, Email [email protected]; [email protected]
Purpose: Cold seawater immersion aggravates hemorrhagic shock-induced homeostasis imbalance and organ dysfunction, leading to increased mortality. Previous studies have shown that treatments targeting oxidative stress and mitochondrial dysfunction have limited efficacy for cold seawater immersion combined with hemorrhagic shock (SIHS). Thus, the mechanisms responsible for SIHS need further investigation.
Methods and Results: Data from the hemorrhagic shock transcriptome and cold seawater immersion targets used for bioinformatics analysis revealed the involvement of endoplasmic reticulum stress (ERS) in SIHS occurrence and progression. Based on these findings, the effects and possible mechanism of inhibiting ERS in SIHS rats were investigated. SIHS causes a lethal triad and impairment of vital organ function, leading to death. Compared to lactated Ringer’s solution, the ERS inhibitor 4-phenylbutyric acid (PBA)significantly ameliorated acidosis and coagulopathy and protected vital organ function while prolonging survival and the golden treatment time. Through target screening and validation, 7 targets were identified for the ERS inhibitor PBA for the treatment of SIHS, among which S1PR1, MMP8 and CFTR may play more important roles.
Conclusion: ERS plays a crucial role in the progression of SIHS. Inhibition of ERS caused by SIHS alleviates the lethal triad, protects organ function, and prolongs survival and the golden treatment time. The ERS inhibitor PBA may be an effective therapeutic measure for treating SIHS.
Keywords: cold seawater immersion, hemorrhagic shock, lethal triad, organ function, ER stress, 4-phenylbutyric acid
Introduction
With the increasing frequency of marine development activities and local wars at sea, the treatment of maritime casualties caused by maritime conflicts and operational accidents has become a challenge worldwide. When victims of a marine accident suffer from hemorrhagic shock, a lower core body temperature, acidosis, and coagulation dysfunction are observed, which is referred to as the “lethal triad”.1,2 Conventional treatments, including fluid resuscitation and the use of adrenaline, cannot achieve target effects. Hence, it is of utmost importance to look for specific measures to recover, ameliorate the lethal triad and organ dysfunction, and prolong the golden window for marine accident victims.
The endoplasmic reticulum (ER) is a crucial organelle for protein synthesis, folding, and modification. The protein folding function of the ER is impaired when it is subjected to endogenous or exogenous stimuli such as hypoxia, oxidative stress, lipid overload, abnormal calcium levels, and infection. The accumulation of misfolded or unfolded proteins in the ER can disrupt protein homeostasis and trigger a series of adaptive responses known as endoplasmic reticulum stress (ERS).3,4 Excessive ERS can lead to several pathological processes, including cell death.5 Previous studies have shown that pathogenic factors such as hypothermia, seawater aspiration and hemorrhagic shock lead to ERS.6–8 However, it is unclear whether ERS plays a role in cold seawater immersion combined with hemorrhagic shock (SIHS) and whether the inhibition of ERS could be a new preventive and therapeutic measure following SIHS.
4-Phenylbutyric acid (PBA), a specific ERS inhibitor that can stabilize peptide structure and enhance endoplasmic reticulum processing of folded proteins, has been used to treat various diseases, such as diabetes and cystic fibrosis.9 In addition, our previous study showed that PBA has anti-inflammatory, antioxidant, immune response modulatory, and organ protective effects via ERS inhibition in septic rats.10,11 It is unclear whether PBA can improve the lethal triad and organ dysfunction in SIHS and extend its golden window of therapeutic.
The aim of the present study was to investigate the effects of PBA on SIHS and explore its possible mechanisms. Models of uncontrolled hemorrhagic shock combined with 15°C seawater immersion in rats were constructed to assess the effects of PBA on the lethal triad, organ function, survival and the golden treatment time. Network pharmacology was used to explore the mechanism of PBA.
Material and Methods
Ethical Statement
All animal experiments were performed in strict accordance with the principles of the Guide for the Care and Use of Laboratory Animals (eighth edition, 2011, National Academies Press, Washington, DC) and approved by the Laboratory Animal Welfare and Ethics Committee of the Army Medical Center, Army Medical University (license No. SCXK [Yu]20170002). None of the authors are members of this committee.
Animal Model and Experimental Protocols
The animals were randomly distributed into the following groups:
Male Sprague Dawley (SD) rats (200 ± 20 g) were fed at the animal care center of Army Medical Center, Army Medical University (license No. SCXK [Yu]20170002). Rats were fasted overnight but allowed water freely before the experiments. On the day of the experiment, the rats were anesthetized with sodium pentobarbital (30 mg/kg) to ensure that they did not respond to needle stimulation but were able to breathe spontaneously without mechanical ventilation. The carotid artery and jugular vein were catheterized with a polyethylene catheter (outer diameter, 0.965 mm; inner diameter, 0.58 mm), and a temperature probe was placed adjacent to the carotid artery for core body temperature monitoring. Carotid artery intubation was used for monitoring blood pressure with an electronic sphygmomanometer (−500-500 KPA/0.4, HaoGan, China) and blood extraction, and jugular vein intubation was used for drug administration. To prevent clotting, the carotid artery catheter and jugular vein catheter were filled with normal (0.9%) saline containing 30 U/mL heparin. A model of shock with persistent hemorrhage was induced by transection of the splenic parenchyma and one of the branches of the splenic artery and vein, as described previously by our research team.13 Blood was allowed to flow freely into the abdominal cavity. Then, the rats were vertically immersed in seawater at 15°C, with armpits above the surface to maintain normal breathing. When the MAP decreased to 40 mmHg (90–120 min), the model was established for subsequent experiments.
The experiment was performed according to the following protocol (Figure 1). In the first part, SIHS targets were obtained and used for enrichment analysis to identify relevant pathways. In the second part, the rats were immediately placed in a 43°C incubator to rewarm and maintain a core body temperature of 34°C after the model was finished. At the same time, the rats were resuscitated with permissive hypotensive resuscitation for 1 hour with LR, and the rats in the PBA groups were administered 5 or 20 mg/kg PBA. Subsequently, the splenic artery was completely ligated to stop the bleeding, and then the rats were resuscitated with LR (to 2 × blood loss) through a jugular vein catheter (25 mL/h) in a room temperature environment. The parameters were measured at the designated time points (Figure 1B). (The resuscitation solution was bathed in water to 37°). In the third part, to observe the effects of PBA on prolonging golden treatment time, the rats did not receive ligation after SIHS. LR or PBA was infused uninterrupted according to the design MAP at 50–60 mmHg until the MAP decreased to 20 mmHg. In the fourth part, the network pharmacological approach was developed to identify the targets of PBA for the treatment of SIHS and validated by molecular docking and RT‒qPCR.
 |
Figure 1 Experimental protocol. (A) Target screening and enrichment analysis of the cold seawater immersion combined with hemorrhagic shock. (B) Effects of PBA on Lethal triad, vital organ function and survival when the splenic artery was ligated after fluid resuscitation for 1 h. (C) Effects of PBA administration on tolerance time during the permissive hypotensive resuscitation period following the model. (D) Workflow of exploration of the mechanism of PBA in the treatment of SIHS based on network pharmacology and molecular docking. Phase I was the establishment of model of cold seawater immersion combined with hemorrhagic shock. Phase II was the permissive hypotensive resuscitation period. Phase III was the observation period. |
Parametric Measurement
Core Temperature and Blood Gas
Core temperature and blood gas were measured by a physiological recorder (SP844) and a blood gas analyzer (ABL800), respectively. Blood gases, including hydrogen potential (pH), lactic acid (Lac), bicarbonate (HCO3-) and alkali surplus (BE), were measured.
Coagulation Parameters and Vital Organ Function Measurements
We collected 2 mL of arterial blood (sodium citrate: blood = 1:9) and centrifuged it for 15 min at 1500 g to measure coagulation function (including activated partial prothrombin time [APTT], prothrombin time [PT], fibrinogen [FIB], international normalized ratio of prothrombin time [PT-INR]) using the ACL TOP 700 system and 3 mL of arterial blood sample for the determination of cardiac, liver and renal function (including cardiac function troponin T [cTnT], aspartate aminotransferase [AST], alanine aminotransferase [ALT], blood urea nitrogen [BUN], and serum creatinine [Scr]) by a biochemical analyzer (DX800, Beckman, Fullerton, CA). One milliliter of arterial blood was used to detect D-lactate, reflecting the function of the intestinal barrier, which was determined by an enzymatic technique.
Blood Loss, Infusion Volume and Animal Survival
At the end of Phase II, the amount of blood loss in the rat abdominal cavity was collected and measured using sterile cotton. The volume of fluid infusion during Phases II and III was also noted. At the end of Phase III, the catheters were removed, the incisions were closed, and the rats were returned to their cages and singly housed for 24 h of monitoring. Mortality was defined by no heartbeat or absence of respiration, and rats that survived for 24 h were euthanized by an overdose of sodium pentobarbital.
Database Analysis of Publicly Available Datasets
Microarray Data and Differential Expression Analysis
The hemorrhagic shock-associated dataset GSE64711 and an annotation file for GPL19607 were downloaded from the Gene Expression Omnibus (GEO).15 A total of 172 samples from patients with hemorrhagic shock and 11 samples from normal subjects were obtained by screening of samples from young males (18–45 years old). Principal component analysis (PCA) was performed using the R package “factoextra”, heatmap clustering analysis was performed using the “pheatmap” package, and differential expression analysis was performed using the “limma” package. Genes with the following criteria were considered differentially expressed genes (DEGs): (1) a |log2 (fold-change)| >1 and (2) an adjusted P < 0.05.
Screening of Cold Seawater Immersion Targets
The NCBI gene database, GeneCards database, and OMIM database were used to search for targets by using “(seawater immersion) OR cold-water immersion” as the keyword. In the NCBI Gene Database, the GeneCards, OMIM and Human Phenotype Ontology Database (https://hpo.jax.org/app/) were used to search for targets by using “hypothermia” as the keyword. The above obtained gene targets were combined and de-duplicated as cold seawater immersion targets. Then, the CTDbase, SEA, STITCH, and ChemMapper databases were used to screen the predicted target proteins of PBA.
Construction of the protein‒protein Interaction (PPI) Network and Core Target Selection
PPI network analysis was conducted using STRING, which is an online database of known and predicted protein‒protein interactions. To clarify the core targets of disease, we used the STRING platform to build the interactions between the DEGs of shock and low-temperature seawater immersion targets and used Cytoscape V3.7.1 to calculate the degree values, betweenness values and MCODE scores of the PPI networks to select the core targets. Cytoscape and Venny 2.1 were utilized to visualize the network and for Venn target mapping, respectively. The same methods were used for PBA and disease target mapping and PPI core target selection.
Enrichment Analysis and Construction of the “Drug-Target-Disease-Pathway” Network
To analyze the main functions of the targets, the clusterProfiler package of R software was used to analyze Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways, determine the significance (P<0.05), and draw the bubble chart. The online website was used to visualize the enrichment results. The common targets of PBA and disease and their KEGG signaling pathways were imported into Cytoscape 3.7.2 to construct a “PBA-target - disease - pathway” network.
In silico Target Validation by Molecular Docking Simulation
CB-DOCK2 integrates cavity detection and a template-based blind docking module, which enables users to obtain potential binding sites and binding modes by referring to known protein–ligand structure information.17 To validate important targets of PBA, we used CB-DOCK2 to perform molecular virtual docking. The pdb files of the protein targets from the RCSB PDB database were collected and treated with PyMOL for dehydration and removal of small-molecule ligands. The structure files of PBA were uploaded in mol2 format, and docking simulation was subsequently performed at CB-DOCK2.
Statistical Analysis
The data are presented as the mean ± SD of n observations. The significant differences among the groups were analyzed by one-way or two-way ANOVA, followed by the post hoc Tukey’s test for multiple comparisons between two groups. The animal survival time was analyzed by median and interquartile ranges. Data processing was performed with the SPSS 20.0 statistical analysis software package. A p value less than 0.05 was considered significant.
Results
ERS Might Play an Important Role in Organ Dysfunction Following Cold Seawater Immersion Combined with Hemorrhagic Shock
To observe the mechanism of injury in SIHS, the hemorrhagic shock dataset GSE64711 (man and young adults [18–45 years old]) and cold seawater immersion-related targets were acquired for analysis. The results identified 332 differentially expressed genes (DEGs) in the shock group compared to the normal control group, including 172 upregulated genes and 160 downregulated genes (Figure 2A–C). At the same time, screened cold seawater immersion-related targets were combined with shock DEGs both as SIHS targets. Cytoscape was used to analyze the network topology of the targets (including Degree value, Betweenness value and MCODE score), and the 129 targets of the diseases were obtained by taking the intersection after screening the targets with the median as the cutoff value (Figure 2D and E).
 |
Figure 2 Target screening and bioinformatics analysis of the cold seawater immersion combined with hemorrhagic shock. (A–C) Heat map, principal component analysis and volcano diagram of the expression matrix for hemorrhagic shock. (D) Databases used to obtain targets related to cold seawater immersion. (E) Venn diagram of all targets (cold seawater immersion-related targets and DEGs of hemorrhagic shock) based on three topological analysis algorithms. (F) Functional enrichment diagram of disease targets. |
GO analysis revealed that the disease targets were associated with important biological processes, including response to decreased oxygen levels, cytokine secretion, intrinsic apoptotic signaling pathway in response to endoplasmic reticulum stress, response to endoplasmic reticulum stress, rough endoplasmic reticulum, endoplasmic reticulum lumen, NADPH oxidase complex, and oxidoreductase complex (Figure 2F). It is evident that endoplasmic reticulum stress plays an important role in SIHS, providing clues for further research.
Protective Effect of PBA on the Lethal Triad Following Cold Seawater Immersion Combined with Hemorrhagic Shock in Rats
The lethal triad is a key characteristic of SIHS and is a major cause of organ dysfunction. To observe the effects of ERS inhibition on the lethal triad, the parameters were measured at the designated time points.
Changes in Acidosis
Acidosis resulting from the accumulation of acid in the body is an important cause of organ dysfunction.18 The rats showed severe acidosis after SIHS, as reflected by an increase in lactate to 13.38 mmol/L and a decrease in pH to 7.04. There was no significant change in LR solution alone group, whereas lactate in the PBA group (5 and 20 mg/kg) significantly decreased to 8.56 and 6.1 mmol/L, respectively, and increased the pH to 7.19 and 7.29, respectively (Figure 3A–D). In addition, the intergroup values of HCO3- and BE exhibited similar trends with respect to pH.
 |
Figure 3 Effects of PBA on the lethal triad following cold seawater immersion combined with hemorrhagic shock. (A–D) Blood gas; pH: Potential of hydrogen, Lac: lactic acid, HCO3-: bicarbonate, BE: alkali surplus. (E–H) Coagulation Function; APTT: activated partial prothrombin time, PT: prothrombin time, FIB: fibrinogen, PT-INR: international normalized ratio of prothrombin time. (I)Core temperature. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001. |
Changes in Coagulation Function
The rats presented coagulation disturbances after SIHS, including increases in APTT, PT, and PT-INR and a decrease in FIB, which could not be reversed by LR resuscitation alone (Figure 3E–H). The APTT, PT and PT-INR in the PBA resuscitation group (5 mg/kg) decreased to 33.04 s, 14.36 s, and 1.25, respectively, and the FIB increased to 1.01 mg/mL; however, a higher concentration of PBA (20 mg/kg) failed to further improve coagulation dysfunction (Figure 3E–H). The results indicated that the ERS inhibitor PBA was beneficial to the improvement of coagulation dysfunction after SIHS, but there was no significant difference between the two concentration groups.
Changes in Body Temperature
The core body temperature of the rats decreased to approximately 33°C after SIHS, and rats without emergency treatment died within 30 min, with further decreases in core body temperature. In the present study, a stepwise rewarming protocol was adopted according to our previous study. Rats were rewarmed to 34°C in the incubator for 1 h and then transferred to a room temperature environment for further rewarming, and the core body temperature of the rats was maintained at approximately 37°C (Figure 3I). All rats could maintain their designed core body temperature.
PBA Played a Protective Role in Vital Organ Function Following Cold Seawater Immersion Combined with Hemorrhagic Shock in Rats
Vital organ dysfunction is a critical factor in the occurrence and progression of hemorrhagic shock and is the major cause of death in shock patients.19 Damage indicators of the heart, liver, kidneys and intestines were compared between the groups. The results showed that the functions of the liver, kidney, heart and intestines were significantly impaired after SIHS in rats, and the indices of liver damage (ie, AST and ALT), kidney damage (BUN and Crea), heart damage (ie, cTnT) and intestinal damage (D-Lac) were significantly increased to 373.50 U/L, 170.23 U/L, 13.11 mmol/L, 54.86 mmol/L, 5.48 μg/L and 2.01 mmol/L, respectively (Figure 4A–F). The damage to organ function could not be alleviated by the LR solution alone but could be significantly improved by PBA, especially ALT, cTnT, and D-Lac, which were reduced to 62.30 U/L, 1.79 μg/L, and 1.28 mmol/L, respectively, in the 20 mg/kg PBA group.
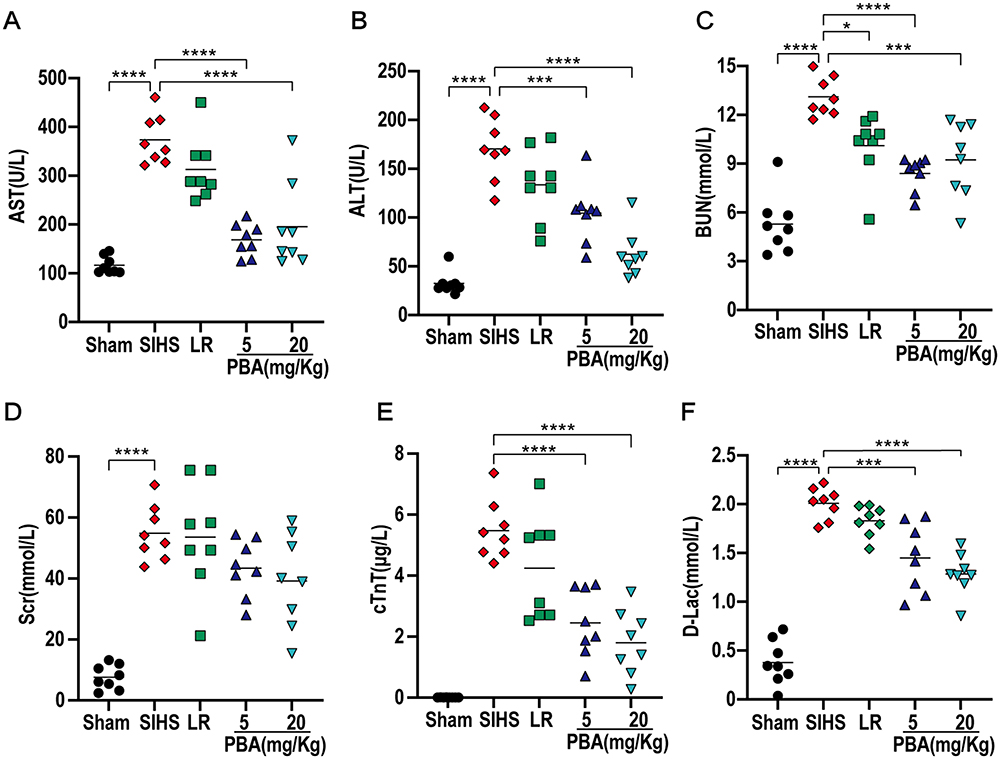 |
Figure 4 Effects of PBA on vital organ functions following cold seawater immersion combined with hemorrhagic shock. (A) AST: aspartate aminotransferase; (B) ALT: alanine aminotransferase; (C) BUN: blood urea nitrogen; (D) Scr: serum creatinine; (E) cTnT: cardiac troponin T; (F) D-Lac: D-lactate. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001. |
PBA Prolonged the Survival of Rats After Cold Seawater Immersion Combined with Hemorrhagic Shock
Without resuscitation following SIHS, the blood loss rate was 30.57% ± 6.59%, which increased to 46.14% ± 8.37% after LR. PBA administration relieved LR-induced blood loss, and the blood loss rates were 36.81% ± 6.38% and 36.92% ± 5.45% in the 5 mg/kg PBA group and 20 mg/kg PBA group, respectively. The results showed that the volume of resuscitation decreased with the decrease of blood loss (Resuscitate twice the amount of blood loss). The volume of resuscitation in the PBA group was significantly lower than that in the LR group, but there was no significant difference between the two concentrations. (Figure 5A and B). Only 25% (4/16) of the rats in the LR group survived for 6 h, and the average survival time was 4.22 h ± 1.83 h. The survival time and survival rate of the rats in the PBA group were significantly greater than those in the LR alone group. The survival time of the 5 mg/kg PBA group was 6.64 h ± 5.01 h, which was prolonged to 11.86 h ± 8.99 h in the 20 mg/kg PBA group (Figure 5C and D).
 |
Figure 5 Effects of PBA on blood loss and survival following cold seawater immersion combined with hemorrhagic shock. (A) Blood loss volume; (B) Fluid requirement; (C) Survival time; (D) Survival rate. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001. |
PBA Prolonged the Golden Treatment Time for Rats Subjected to Cold Seawater Immersion Combined with Hemorrhagic Shock
The evacuation time after trauma is commonly 1 h, which is considered the golden treatment time. However, 1 h of gold treatment time is not enough under harsh conditions. It is not clear whether PBA can buy golden treatment time in SIHS.
Consistent with expectations, the MAP decreased sharply without resuscitation. The average hypotensive maintenance time was 78 ± 30 min in the LR group, and the administration of PBA (5 or 20 mg/kg) significantly prolonged the hypotensive maintenance time to 135 min ± 45 min and 153 min ± 65 min, respectively (Figure 6A and B). The blood loss volume was 55.52%±6.61% with LR resuscitation alone, and PBA (5 or 20 mg/kg) significantly reduced blood loss to 40.96%±4.81% and 42.45%±6.19%, respectively (Figure 6C). Correspondingly, the PBA also significantly reduced the infusion volume by more than 10 mL compared to that in the LR group (Figure 6D).
 |
Figure 6 PBA prolonged the golden treatment time for the cold seawater immersion combined with hemorrhagic shock. (A) Mean arterial pressure (MAP). (B) The hypotensive maintenance time of LR or PBA. (C and D) Blood loss and infusion volume in permissive hypotensive maintenance period. **P<0.01, ***P<0.001, ****P<0.0001. |
The Possible Mechanism of PBA in the Protection of Cold Seawater Immersion Combined with Hemorrhagic Shock
To investigate the mechanism of the endoplasmic reticulum stress inhibitor PBA in SIHS, network pharmacology and molecular docking were used. A total of 404 PBA-related targets were obtained via the CTD, SEA, STITCH and Chemmapper drug target databases by using “4-phenylbutyric acid” as the keyword. The disease targets related to SIHS were intersected with PBA-related targets to obtain 33 common targets (Figure 7A and B). KEGG pathway enrichment analysis of 33 targets revealed that they were mainly enriched in the HIF-1 signaling pathway, NOD-like receptor signaling pathway, lipid and atherosclerosis pathway, IL-17 signaling pathway, C-type lectin receptor signaling pathway, NF-kappa B signaling pathway, Toll-like receptor signaling pathway, TNF signaling pathway, FoxO signaling pathway and apoptosis pathway, which are closely associated with the immune response and coagulation response. The “PBA-target-disease-pathway” pathway was further analyzed by Cytoscape (3.7.2), and 10 key genes, including IL1B, IL6, S1PR1, MMP8, GPT, MAPK1, PTGS2, PTGER4, CXCL8, and CFTR, were identified (Figure 7C and D).
 |
Figure 7 The key therapeutic targets and pathways of PBA in the treatment of cold seawater immersion combined with hemorrhagic shock (A) PBA chemical structure formula; (B) Venn mapping of disease and drug targets; (C) “PBA-target-disease-pathway” network; (D) Topological scoring of common targets Venn intersection; (E and F) The binding energy of PBA to key target proteins; C1-C5 are the predicted target protein pockets, with lower scores representing higher affinity for the receptor and ligand. |
To further identify the target gene of PBA, molecular docking of key targets to PBA was performed by CB-DOCK2. The Vina scores for the optimal binding modes of PBA to the 10 key target proteins were < −5.0 kcal·mol-1, indicating that the receptors and ligands have good binding properties (Figure 7E and F). PBA has the highest docking affinity for S1PR1 and MMP8, as shown in the molecular docking pattern diagram below (Figure 8).
 |
Figure 8 Molecular models of PBA binding to the predicted target. (A) IL1β; (B) IL6; (C) S1PR1; (D) MMP8; (E) GPT; (F) MAPK1; (G) PTGS2; (H) PTGER4; (I) CXCL8; (J) CFTR. |
Validation of Key Targets of PBA
The expression profiles of key targets were estimated by RT‒qPCR of blood samples following PBA administration. As shown in Figure 9, the mRNA expression of Il1β, Il6, Il8, S1pr1, Mmp8, Gpt and Ptgs2 increased after SIHS, and LR infusion alone decreased Gpt and even increased Mmp8 and Mapk1 expression. PBA administration attenuated the increases in Il1β, Il6, S1pr1, Mmp8, Gpt and Mapk1. In contrast, Ctfr was reduced after SIHS and LR infusion and recovered after PBA administration. Moreover, PBA and LR infusion did not alter the expression of Ptgs2, Ptger4, or Il8. The high concentration (20mg/Kg) of PBA further reduced the mRNA level of IL6 compared to the low concentration group (5mg/Kg). These results suggest that PBA may protect against SIHS by reducing the expression of damage factors, including Il1β, Il6, S1pr1, Mmp8, Gpt, and Mapk1, and restoring Ctfr expression. Combined with the Vina score of molecular docking, it is clear that S1PR1, MMP8 and CFTR may play more important roles in this process.
 |
Figure 9 mRNA expression levels of ten key targets between groups (A) Il1β; (B) Il6; (C) S1pr1; (D) Mmp8; (E) Gpt; (F) Mapk1; (G) Ptgs2; (H) Ptger4; (I) Il8; (J) Cftr. |
Discussion
Prolonged immersion in seawater can lead to a rapid decrease in core body temperature. Hypothermia inhibits the coagulation process, decreasing the activity of coagulation factors and increasing the disruption and adhesion of platelets. Hemorrhage may cause ischemia, hypoxia and metabolic reprogramming. Extensive loss of body fluids and changes in metabolic patterns lead to lactic acid accumulation and metabolic acidosis, which can further worsen this coagulation dysfunction. Thus, hemorrhagic shock accompanied by immersion in seawater often triggers more severe lethal triad, more deteriorative organ dysfunction and greater mortality than that on land. Although rewarming, fluid resuscitation, correction of acidosis and coagulopathy can partly attenuate symptoms in patients with SIHS, the survival rate of patients with SIHS is unsatisfactory. However, many complex pathophysiological mechanisms are still unknown. Thus, further understanding of the mechanisms underlying the pathogenesis of SIHS and exploring novel treatments for SIHS patients are critical. Shock and hypothermia lead to severe hypoxia in the body, and cells are subjected to severe stress triggering ER stress.6,8 In the present study, we used a dataset from shock and cold seawater immersion to screen 129 DEGs related to the SIHS. Both BP and CC analyses showed that multiple ER stress-related signaling pathways were markedly activated. PBA, a low-molecular-weight fatty acid with multiple biological activities, including ER stress inhibition, antioxidative effects, ammonia scavenging and histone deacetylase inhibition, has been shown to have protective effects on various diseases, such as asthma, diabetes and spontaneous hypertension.20–22
Our study demonstrated that the application of PBA has a beneficial effect on SIHS by improving the lethal triad (except for core body temperature) and vital organ function and prolonging the golden treatment time and 24 h survival rate (20 mg/kg of PBA was shown to have better beneficial effects). We presume that the main mechanism underlying the protective effect of PBA on SIHS is the inhibition of ER stress and the reduction of inflammatory factor expression. It has been shown that both shock and cold seawater immersion cause cellular hypoxia to activate ERS, which can fragment mitochondria by promoting the ubiquitination degradation of mitochondrial fusion factor mitofusin-2 (MFN2), increasing the S616/S637 phosphorylation ratio of dynamin-related protein 1 (Drp1) to fragment mitochondria, resulting in a reduction of the mitochondrial matrix and the inner mitochondrial membrane (sites of the tricarboxylic acid cycle and oxidative phosphorylation).23,24 At the same time, ERS-induced mitochondrial calcium overload is involved in the destruction of electron transport chain.25 The above factors conspire to prevent the tricarboxylic acid cycle from continuing, leading to increased glycolysis and lactic acid accumulation. PBA may restore the tricarboxylic acid cycle and oxidative phosphorylation by improving mitochondrial dysfunction through inhibition of ERS and alleviating acidosis. Correction of acidosis in concert with rewarming may also play a great role in the improvement of prothrombin activity and platelet function. Furthermore, the restoration of organ function and survival, especially ALT, cTnT, and D-Lac, by the LR solution with the addition of PBA may also be associated with improvement in the ERS-associated lethal triad and a reduction in the expression level of inflammatory factors. In the part of the effect observation of prolonging the gold treatment time, we also found that the LR solution with the addition of PBA had less blood loss and resuscitation volume than the LR solution alone, and the hypotensive maintenance time was longer than the LR solution alone. On the one hand, PBA can correct coagulopathy and reduce continuous blood loss, thereby prolonging the maintenance time of hypotension. On the other hand, PBA improves cardiovascular function and blood pressure indirectly by improving the function of other organs.
The ER is an intracellular membranous organelle involved in protein synthesis, folding, maturation and posttranslational modification. Therefore, the ER is closely related to intracellular homeostasis, health maintenance and disease pathogenesis. ER stress occurs when unfolded/misfolded proteins accumulate after disturbance of the ER environment to help cells cope with stress.26 In recent years, a large number of studies have reported that ER stress is a novel mechanism for the development and progression of various diseases. It plays an important role in calcium homeostasis, intracellular redox potential and the synthesis and folding of proteins.27 Various factors, including hypoxia, ischemia and infections, may trigger ER stress. The principal function of PBA is to inhibit ER stress as a chemical chaperone.20 Previous studies in our laboratory and others have demonstrated that PBA improves decreased vascular reactivity and protects vascular function by inhibiting antioxidative stress.1128,29 In addition to oxidative stress, whether there are other mechanism that regulate ER stress in SIHS. Our present data suggested that core gene selection and molecular docking simulation showed that PBA interacts with many inflammatory genes. These results indicate that PBA may play a therapeutic role in SIHS by attenuating the expression of inflammatory factors in addition to inhibiting ERS.
The immune status and inflammation level of the body are closely related to the development and prognosis of the disease. S1PR1 is a G protein-coupled receptor involved in regulating the recruitment of immune cells. S1PR1 binds to its ligand sphingosine-1-phosphate (S1P), activates downstream signaling pathways, and thereby promotes the migration and localization of lymphocytes and neutrophils to infected or injured organs, participating in pathogen clearance and inflammation regulation.30,31 Intervening in the S1P/S1PR1 axis can block the NFκB/IL-6/STAT3 signaling cascade, leading to the resolution of inflammation and the induction of neutrophil apoptosis.32 The level of MMP8, a collagenase that significantly increases in inflammatory diseases, leads to collagen degradation, epithelial damage, increased permeability, and neutrophil infiltration.33,34 Our research findings revealed that PBA treatment reduced the expression of S1PR1 and MMP8. The attenuation of the S1P/S1PR1 signaling pathway may result in a decrease in inflammatory factors and prevent the migration, activation, and proliferation of immune cells such as neutrophils after SIHS. The core targets screened in this study included four classic inflammation-related genes, namely, IL-1β, IL-6 and MAPK1. In models of acute lung injury and arthritis, PBA reduces the release of the proinflammatory mediators IL-1β, TNF-α, and IL-6.35–37 Neeraj noted that PBA downregulates the relative transcription levels of Cox-2 and IL-6 in cystic fibrosis by inhibiting the activity of histone deacetylases (HDACs).38 The occurrent study revealed that PBA can reduce the expression of IL-1β, IL-6 and MAPK1 in a rat model of SIHS, consistent with previous research.
The protease Gpt encodes cytosolic alanine amino transaminase 1 (ALT1), whose serum activity is routinely used as a biomarker for drug toxicity and infection-induced liver injury. Several studies have shown that PBA can significantly improve ER stress in acute liver failure or drug-induced liver injury, decrease Gpt expression, and reduce liver dysfunction.39,40 We observed that Gpt increased after SIHS and decreased after PBA treatment with the recovery of liver function. CFTR, a widely expressed endoplasmic reticulum (ER) luminal protein, regulates the biosynthesis and transport of several secretory proteins and is a key regulator of epithelial cell function. Several studies have demonstrated that the key ER stress transcription factor ATF6 binds to the CFTR transcriptional promoter, causing a decrease in CFTR mRNA.41,42 Additionally, ER stress reduces the structural stability of the CFTR protein and causes its degradation in the ER.43 Reduced CFTR causes a decrease in intestinal epithelial chloride secretion and a disruption of the intestinal chemical barrier, leading to the translocation of intestinal pathogenic microorganisms and toxins into the bloodstream and triggering gut-derived infection.12 PBA was shown to increase the expression of CFTR transcripts and assist in CFTR processing, maturation and apical membrane localization by inhibiting ER stress, thereby restoring normal intestinal structural function.14,16 In the present study, PBA could restore the expression of CFTR in SIHS rats, which may also be related to the decrease in the intestinal injury marker D-Lac. PBA elicits organ function protection mainly through the inhibition of inflammation and ER stress.
Although the current study revealed that the PBA was appropriate for the treatment of SIHS, there were still some limitations. First, PBA extends the golden rescue time, except for inhibiting ERS and improving organ function, it is still unclear whether there is also an involvement of hemodynamic changes. Second, this study revealed different effects of PBA on the liver, intestine and kidney, but the mechanism is not clear. Third, the mechanisms by which PBA suppresses inflammation need further exploration. Fourth, the crucial elements among the targets regulated by PBA in SIHS need further investigation.
Conclusion
Cold seawater immersion combined with hemorrhagic shock can increase the mortality of the wounded, conventional resuscitation measures are not effective in treating this condition. ERS inhibitors have multiple organ protection for SIHS animals, the specific mechanism may be related to S1PR1, MMP8 and CFTR.
Abbreviations
SIHS, Cold seawater immersion combined with hemorrhagic shock; ER, endoplasmic reticulum; ERS, endoplasmic reticulum stress; PBA, 4-phenylbutyric acid; LR, lactated Ringer’s solution; pH, Potential of hydrogen; Lac, lactic acid; HCO3-, bicarbonate; BE, alkali surplus; APTT, activated partial prothrombin time; PT, prothrombin time; FIB, fibrinogen; PT-INR, international normalized ratio of prothrombin time; cTnT, cardiac function troponin T; AST, aspartate aminotransferase; ALT, alanine aminotransferase; BUN, blood urea nitrogen; Scr, serum creatinine; DEGs, differentially expressed genes; MAP, Mean arterial pressure; IL1B, Interleukin 1B; IL6, Interleukin 6; S1PR1, Sphingosine-1-phosphate receptor 1; MMP8, Matrix metalloproteinase 8; GPT, Glutamate pyruvate transaminase; MAPK1, Mitogen-activated protein kinase 1; PTGS2, Prostaglandin-endoperoxide synthase 2; PTGER4, Prostaglandin E receptor 4; CXCL8, C-X-C motif chemokine ligand 8; CFTR, Cystic fibrosis transmembrane conductance regulator; S1P, Sphingosine-1-phosphate; TNF-α, Tumor necrosis factor-alpha; HDACs, Histone deacetylases.
Institutional Review Board Statement
The animal study was reviewed and approved by the Ethics Review Committee of the Army Medical Center, Army Medical University (license No. SCXK [Yu]20170002). Human data based on the GEO open source data, so there is no ethical issues and other conflicts of interest. And we obtained exempt ethics approval from the ethics committee at the Army Medical Center.
Acknowledgments
We would like to acknowledge the use of publicly available data from GEO, which was originally published in Critical Care by Erin L Vanzant. We are grateful for the Erin L Vanzant for making these valuable data accessible to the scientific community. In addition we thank the Department of Clinical Laboratory, Army Medical Center, for assisting in the measurement of animal coagulation and multiorgan function.
Funding
This research was funded by the High-level Scientific and Technological Innovation Talent Project of the Army and the China National Natural Science Foundation (Nos.82270523).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Yan H, Mao Q, Ma Y, et al. Seawater immersion aggravates burn injury causing severe blood coagulation dysfunction. Biomed Res Int. 2016;2016:9471478. doi:10.1155/2016/9471478
2. Deng H, Zhu Y, Li Q, et al. Effects of seawater immersion on lethal triad and organ function in healthy and hemorrhagic shock rats. J Surg Res. 2023;284:173–185. doi:10.1016/j.jss.2022.11.070
3. So JS. Roles of endoplasmic reticulum stress in immune responses. Mol Cells. 2018;41(8):705–716. doi:10.14348/molcells.2018.0241
4. Chen X, Shi C, He M, Xiong S, Xia X. Endoplasmic reticulum stress: molecular mechanism and therapeutic targets. Signal Transduct Target Ther. 2023;8(1):352. doi:10.1038/s41392-023-01570-w
5. Gao H, He C, Hua R, et al. Endoplasmic reticulum stress of gut enterocyte and intestinal diseases. Front Mol Biosci. 2022;9:817392. doi:10.3389/fmolb.2022.817392
6. Rzechorzek NM, Connick P, Patani R, Selvaraj BT, Chandran S. Hypothermic preconditioning of human cortical neurons requires proteostatic priming. EBioMedicine. 2015;2(6):528–535. doi:10.1016/j.ebiom.2015.04.004
7. Zhang M, Gao Y, Zhao W, Yu G, Jin F. ACE-2/ANG1-7 ameliorates ER stress-induced apoptosis in seawater aspiration-induced acute lung injury. Am J Physiol Lung Cell Mol Physiol. 2018;315(6):L1015–L1027. doi:10.1152/ajplung.00163.2018
8. Obert DP, Wolpert AK, Korff S. Modulation of endoplasmic reticulum stress influences ischemia-reperfusion injury after hemorrhagic shock. Shock. 2019;52(5):e76–e84. doi:10.1097/SHK.0000000000001298
9. Jeon JH, Im S, Kim HS, et al. Chemical chaperones to inhibit endoplasmic reticulum stress: Implications in diseases. Drug Des Devel Ther. 2022;16:4385–4397. doi:10.2147/DDDT.S393816
10. Liu L, Wu H, Zang J, et al. 4-Phenylbutyric acid reveals good beneficial effects on vital organ function via anti-endoplasmic reticulum stress in septic rats. Crit Care Med. 2016;44(8):e689–701. doi:10.1097/CCM.0000000000001662
11. Kuang L, Zhu Y, Wu Y, et al. Synergetic effect of 4-phenylbutyric acid in combination with cyclosporine a on cardiovascular function in sepsis rats via inhibition of endoplasmic reticulum stress and mitochondrial permeability transition pore opening. Front Pharmacol. 2021;12:770558. doi:10.3389/fphar.2021.770558
12. Fiorotto R, Strazzabosco M. Pathophysiology of cystic fibrosis liver disease: A channelopathy leading to alterations in innate immunity and in microbiota. Cell Mol Gastroenterol Hepatol. 2019;8(2):197–207. doi:10.1016/j.jcmgh.2019.04.013
13. Li T, Zhu Y, Fang Y, Liu L. Determination of the optimal mean arterial pressure for post bleeding resuscitation after hemorrhagic shock in rats. Anesthesiology. 2012;116(1):103–112. doi:10.1097/ALN.0b013e31823de24f
14. Poothong J, Jang I, Kaufman RJ. Defects in protein folding and/or quality control cause protein aggregation in the endoplasmic reticulum. Prog Mol Subcell Biol. 2021;59:115–143. doi:10.1007/978-3-030-67696-4_6
15. Vanzant EL, Hilton RE, Lopez CM, et al. Advanced age is associated with worsened outcomes and a unique genomic response in severely injured patients with hemorrhagic shock. Crit Care. 2015;19(1):77. doi:10.1186/s13054-015-0788-x
16. Brecker M, Khakhina S, Schubert TJ, Thompson Z, Rubenstein RC. The probable, possible, and novel functions of ERp29. Front Physiol. 2020;11:574339. doi:10.3389/fphys.2020.574339
17. Liu Y, Yang X, Gan J, Chen S, Xiao ZX, Cao Y. CB-Dock2: Improved protein-ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic Acids Res. 2022;50(W1):W159–W164. doi:10.1093/nar/gkac394
18. Ahmed AR, Satti MM, Abdalla AE, Giblin L, Lappin D. The prevalence and management of metabolic acidosis of chronic kidney disease. Ir Med J. 2019;112(9):1002.
19. Torres Filho I. Hemorrhagic shock and the microvasculature. Compr Physiol. 2017;8(1):61–101. doi:10.1002/cphy.c170006
20. Delmotte P, Yap JQ, Dasgupta D, Sieck GC. Chemical Chaperone 4-PBA Mitigates tumor necrosis factor alpha-induced endoplasmic reticulum stress in human airway smooth muscle. Int J Mol Sci. 2023;24(21):15816. doi:10.3390/ijms242115816
21. Reddy SS, Shruthi K, Joy D, Reddy GB. 4-PBA prevents diabetic muscle atrophy in rats by modulating ER stress response and ubiquitin-proteasome system. Chem Biol Interact. 2019;306:70–77. doi:10.1016/j.cbi.2019.04.009
22. Zhu J, Shao A, Wang C, Zeng C, Wang H. Inhibition of endoplasmic reticulum stress restores the balance of renal RAS components and lowers blood pressure in the spontaneously hypertensive rats. Clin Exp Hypertens. 2023;45(1):2202367. doi:10.1080/10641963.2023.2202367
23. Dasgupta D, Delmotte P, Mahadev Bhat S, et al. TNFα Mediated Endoplasmic Reticulum Stress Promotes Differential Phosphorylation of DRP1 and Mitochondrial Fragmentation in Human Airway Smooth Muscle Cells. The FASEB Journal. 2022;36:. doi:10.1096/fasebj.2022.36.s1.r3133
24. Chen Y, Dorn GW. PINK1-phosphorylated mitofusin 2 is a Parkin receptor for culling damaged mitochondria. Science. 2013;340(6131):471–475. doi:10.1126/science.1231031
25. Balderas E, Lee SH, Rai NK, Mollinedo DM, Duron HE, Chaudhuri D. Mitochondrial calcium regulation of cardiac metabolism in health and disease. Physiology. 2024;39(5):1. doi:10.1152/physiol.00014.2024
26. Ren J, Bi Y, Sowers JR, Hetz C, Zhang Y. Endoplasmic reticulum stress and unfolded protein response in cardiovascular diseases. Nat Rev Cardiol. 2021;18(7):499–521. doi:10.1038/s41569-021-00511-w
27. Bonsignore G, Martinotti S, Ranzato E. Endoplasmic reticulum stress and cancer: Could unfolded protein response be a druggable target for cancer therapy? Int J Mol Sci. 2023;24(2):1566. doi:10.3390/ijms24021566
28. Morris HE, Neves KB, Nilsen M, Montezano AC, MacLean MR, Touyz RM. Notch3/Hes5 induces vascular dysfunction in hypoxia-induced pulmonary hypertension through er stress and redox-sensitive pathways. Hypertension. 2023;80(8):1683–1696. doi:10.1161/HYPERTENSIONAHA.122.20449
29. Yang G, Peng X, Hu Y, et al. 4-Phenylbutyrate benefits traumatic hemorrhagic shock in rats by attenuating oxidative stress, not by attenuating endoplasmic reticulum stress. Crit Care Med. 2016;44(7):e477–91. doi:10.1097/CCM.0000000000001469
30. Zhao X, Yang L, Chang N, et al. Neutrophil recruitment mediated by sphingosine 1-phosphate (S1P)/S1P receptors during chronic liver injury. Cell Immunol. 2021;359:104243. doi:10.1016/j.cellimm.2020.104243
31. Chen H, Wang J, Zhang C, et al. Sphingosine 1-phosphate receptor, a new therapeutic direction in different diseases. Biomed Pharmacother. 2022;153:113341. doi:10.1016/j.biopha.2022.113341
32. Perez DA, Galvao I, Athayde RM, et al. Inhibition of the sphingosine-1-phosphate pathway promotes the resolution of neutrophilic inflammation. Eur J Immunol. 2019;49(7):1038–1051. doi:10.1002/eji.201848049
33. Moliere S, Jaulin A, Tomasetto CL, Dali-Youcef N. Roles of matrix metalloproteinases and their natural inhibitors in metabolism: Insights into health and disease. Int J Mol Sci. 2023;24(13):10649. doi:10.3390/ijms241310649
34. Daly MC, Atkinson SJ, Varisco BM, et al. Role of matrix metalloproteinase-8 as a mediator of injury in intestinal ischemia and reperfusion. FASEB J. 2016;30(10):3453–3460. doi:10.1096/fj.201600242R
35. Zeng M, Sang W, Chen S, et al. 4-PBA inhibits LPS-induced inflammation through regulating ER stress and autophagy in acute lung injury models. Toxicol Lett. 2017;271:26–37. doi:10.1016/j.toxlet.2017.02.023
36. Choi Y, Lee EG, Jeong JH, Yoo WH. 4-Phenylbutyric acid, a potent endoplasmic reticulum stress inhibitor, attenuates the severity of collagen-induced arthritis in mice via inhibition of proliferation and inflammatory responses of synovial fibroblasts. Kaohsiung J Med Sci. 2021;37(7):604–615. doi:10.1002/kjm2.12376
37. Jia H, Zhang T, Liu N, et al. 4-Phenylbutyric acid alleviates 3-acetyldeoxynivalenol-induced immune cells response by inhibiting endoplasmic reticulum stress in mouse spleen. Food Chem Toxicol. 2022;164:113002. doi:10.1016/j.fct.2022.113002
38. Vij N, Amoako MO, Mazur S, Zeitlin PL. CHOP transcription factor mediates IL-8 signaling in cystic fibrosis bronchial epithelial cells. Am J Respir Cell Mol Biol. 2008;38(2):176–184. doi:10.1165/rcmb.2007-0197OC
39. Jia H, Liu N, Zhang Y, Wang C, Yang Y, Wu Z. 3-Acetyldeoxynivalenol induces cell death through endoplasmic reticulum stress in mouse liver. Environ Pollut. 2021;286:117238. doi:10.1016/j.envpol.2021.117238
40. Wu D, Yang C, Yang M, et al. Citrinin-Induced Hepatotoxicity in Mice Is Regulated by the Ca(2+)/Endoplasmic Reticulum Stress Signaling Pathway. Toxins. 2022;14(4):259. doi:10.3390/toxins14040259
41. Trouve P, Ferec C, Genin E. The interplay between the unfolded protein response, inflammation and infection in cystic fibrosis. Cells. 2021;10(11):2980. doi:10.3390/cells10112980
42. Pecoraro M, Serra A, Pascale M, Franceschelli S. Vx-809, a CFTR corrector, acts through a general mechanism of protein folding and on the inflammatory process. Int J Mol Sci. 2023;24(4):4252. doi:10.3390/ijms24044252
43. Cui X, Zhang Y, Lu Y, Xiang M. ROS and endoplasmic reticulum stress in pulmonary disease. Front Pharmacol. 2022;13:879204. doi:10.3389/fphar.2022.879204
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.