")
Back to Journals » Journal of Inflammation Research » Volume 17
Role of Pyroptosis in Endometrial Cancer and Its Therapeutic Regulation
Authors Al Mamun A , Geng P, Wang S , Shao C
Received 13 July 2024
Accepted for publication 21 September 2024
Published 3 October 2024 Volume 2024:17 Pages 7037—7056
DOI https://doi.org/10.2147/JIR.S486878
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Abdullah Al Mamun,1,2 Peiwu Geng,1 Shuanghu Wang,1 Chuxiao Shao1
1Key Laboratory of Joint Diagnosis and Treatment of Chronic Liver Disease and Liver Cancer of Lishui, The Lishui Hospital of Wenzhou Medical University, The First Affiliated Hospital of Lishui University, Lishui People’s Hospital, Lishui, Zhejiang, 323000, People’s Republic of China; 2Molecular Pharmacology Research Center, School of Pharmaceutical Sciences, Wenzhou Medical University, Wenzhou, Zhejiang, 325000, People’s Republic of China
Correspondence: Chuxiao Shao; Shuanghu Wang, Key Laboratory of Joint Diagnosis and Treatment of Chronic Liver Disease and Liver Cancer of Lishui, the Lishui Hospital of Wenzhou Medical University, the First Affiliated Hospital of Lishui University, Lishui People’s Hospital, Lishui, Zhejiang, 323000, People’s Republic of China, Email [email protected]; [email protected]
Abstract: Pyroptosis is an inflammatory cell death induced by inflammasomes that release several pro-inflammatory mediators such as interleukin-18 (IL-18) and interleukin-1β (IL-1β). Pyroptosis, a type of programmed cell death, has recently received increased interest both as a therapeutic and immunological mechanism. Numerous studies have provided substantial evidence supporting the involvement of inflammasomes and pyroptosis in a variety of pathological conditions including cancers, nerve damage, inflammatory diseases and metabolic conditions. Researchers have demonstrated that dysregulation of pyroptosis and inflammasomes contribute to the progression of endometriosis and gynecological malignancies. Current research also indicates that inflammasome and pyroptosis-dependent signaling pathways may further induce the progression of endometrial cancer (EC). More specifically, dysregulation of NLR family pyrin domain 3 (NLRP3) and caspase-1-dependent pyroptosis play a contributory role in the pathogenesis and development of EC. Therefore, pyroptosis-regulated protein gasdermin D (GSDMD) may be an independent prognostic biomarker for the detection of EC. This review presents the molecular mechanisms of pyroptosis-dependent signaling pathways and their contributory role and function in advancing EC. Moreover, this review offers new insights into potential future applications and innovative approaches in utilizing pyroptosis to develop effective anti-cancer therapies.
Keywords: endometrial cancer, pyroptosis, NLRP3, caspase-1, GSDMD, IL-1β, IL-18
Graphical Abstract:

Introduction
Endometrial cancer (EC) is the predominant form of gynecologic malignancy worldwide. The incidence of EC is approximately 142,000 women per year with an estimated 42,000 women dying from this disease.1 There is currently a predominant role for surgical interventions in the management of patients without reproductive needs.2 However, patients with elevated risk factors may require additional treatment techniques such as radiation and chemotherapy. These therapeutic approaches promote ROS-mediated stress and induce apoptosis in cancer cells. Cancer-related deficiencies in apoptosis initiation may occur, but there is also the possibility of an “oncogene addiction” phenomenon that can result in treatment ineffectiveness.3 Cancer cells are exquisitely dependent upon a single oncogenic lesion through many genetic and epigenetic changes during the development of their neoplastic characteristics. It was found in a laboratory setting that tumor cells could depend on tumor cells when an oncogene was suppressed or tumor suppressor expression was restored, which led to the initial suggestion that the developemnt of tumor cells could depend on tumor cells. These first discoveries suggested that therapeutic drugs aimed at the repair or regulation of these mutant gene products might have broad effectiveness in the treatment of EC. Apoptosis, necroptosis and pyroptosis are discrete cellular mechanisms of programmed cell death within host cells.4,5 Recent data suggests a significant relationship between pyroptosis and the pathogenesis and progression of multiple diseases including cardiovascular diseases (CVDs),6–9 cancer,10–12 neurological diseases (NDs)13–15 and metabolic diseases (MDs).16–18 In addition, pyroptosis plays a key role in the onset and advancement of several inflammatory conditions, particularly in the conversion of organ or cell inflammation into cancer.
The purpose of this study was to find, screen and evaluate information on inflammasomes and pyroptosis-dependent cell death in EC including information obtained from Elsevier, Google Scholar, PubMed, Science Direct, Scirus, Sci Finder, Scopus, Springer and Web of Science. We also identified potential therapeutic compounds that may be used to suggest and control these mechanisms. Furthermore, we used local and international books and peer-reviewed journals to find relevant information. This review also discusses the molecular mechanisms of pyroptosis-related signaling pathways and their contributory roles in the pathogenesis and progression of EC.
Historical and Biological Features and Functions of Pyroptosis
Pyroptosis originates primarily from the Greek words “pyro” and “ptosis”, which represent fever and falling, correspondingly.4,19 This terminology is employed to describe a newly identified form of programmed cell death (PCD) that has inflammatory characteristics. In 1990, researchers discovered that Shigella flexneri and Salmonella infections remove mouse macrophages or human monocytes.20 Shigella dysenteriae was reported to activate caspase-1 in host cells by Arturo Zychlinsky in 1997.21 Hersh et al also showed that the inhibition of caspase-1 alleviated Salmonella-induced cell death.22 Lawrence H. Boise and Brad Cookson reported in 2001 that microbial infection caused by bacteria led to the death of macrophages via the activation of caspase-1-dependent programmed necrosis.23,24 Pyroptosis and apoptosis have comparable biological features and functions including DNA fragmentation and chromatin. Specifically, pyroptotic cells show denaturation and the development of spherical expansions on their cellular membranes.25 The differences between pyroptosis and apoptosis can be distinguished based on a wide range of characteristics. Pyroptosis is mainly characterized by a rapid disruption of the plasma membrane, as opposed to the blebbing of the membrane that usually occurs during apoptosis. Pyroptosis results in rapid plasma membrane disintegration, whereas apoptosis is comparatively slow.26 The degradation of the membrane structure leads to the infiltration of water and ions into the cell, which ultimately causes the cell to enlarge and burst.27 Pyroptosis also exhibits similar characteristics to apoptosis including caspase activation and PtdSer exposure.28 Pyroptotic cells produce PtdSer, which attracts engulfing cells. However, this mechanism differs from the process of PtdSer exposure mediated by scramblase in apoptosis.29 Studies have shown that pyroptosis results in the externalization of PtdSer through membrane disintegration.30 Nuclear condensation and DNA damage are found in pyroptotic cells, but the nuclei remain undamaged compared to apoptosis. Apoptosis involves the caspase-3 enzyme. Pyroptosis exhibits distinct cellular properties that differentiate it from other types of cell death.31–33 Pyroptosis induces low-grade inflammation, whereas apoptosis does not cause inflammation.34 Pyroptosis is triggered by both external and internal signals such as viral and bacterial infections, being exposed to toxins and certain chemotherapeutic agents.35–37 In contrast to necrosis, pyroptosis involves cytoplasmic damage caused by sudden plasma membrane rupture. Activation of caspases or release of granzymes leads to the oligomerization of gasdermin N-terminal and the development of pores (1–2 μm) in plasma membranes, which further promotes the matured form of IL-1β/IL-18 (4.5 nm) and caspase-1 (7.5 nm) permeability.38 The ingress of water via holes stimulates cellular expansion, osmotic disintegration and plasma membrane rupture, resulting in the release of pro-inflammatory factors including IL-1β and IL-18.39,40 Pyroptotic cell permeability is facilitated by the low molecular weight of 7-amino-actinomycin (7-AAD), phosphatidylinositol (PI) and ethidium bromide (EtBr). Pyroptotic cells eliminate these dyes by maintaining membrane integrity, whereas apoptotic cells destroy their membrane.41 Annexin V is a marker for pyroptotic cells, which are similar to apoptotic cells. It forms a bond with phosphatidyl serine (PS). Consequently, Annexin V cannot distinguish between cells undergoing apoptosis and those undergoing pyroptosis. Apoptotic bodies are generated during the process of apoptosis, whereas pyroptotic bodies are generated during the process of pyroptosis.42 Pyroptotic bodies have a diameter of 1–5 µm, which is comparable to the size of apoptotic bodies.43
Ferroptosis causes a typical- morphological feature with intact cell membranes and nuclei devoid of chromatin condensation. However, mitochondria exhibit reduced cristae and membranes collapse and rupture.44 It is triggered by disrupting the glutathione-dependent antioxidant defense using defects in system XC− or glutathione peroxidase 4 (GPX4). Glutathione (GSH) is synthesized in the cell by converting extracellular cystine into cysteine.45 GPX4 can swiftly facilitate the interaction between glutathione and lipid hydroperoxides, resulting in a decrease in the amount of lipid peroxidation inside cells. Inhibition of GPX4 or depletion of GSH leads to the accumulation of lipid hydroperoxides. It has been demonstrated that unbound iron reacts with lipid hydroperoxides via the Fenton reaction, which further produces reactive oxygen species (ROS) from lipids. The excessive production of ROS causes cell death. Ferroptosis activation may be verified by administering ferroptosis inhibitors (eg, liproxstatin-1 and ferrostatin-1) and by determining lipid peroxides (eg, malondialdehyde quantification and 4-hydroxynonenal quantification).46
Necroptosis, also called programmed necrosis, is featured by activating receptor-interacting protein kinases (RIPKs) via regulating a variety of signaling pathways.47 RIPKs are triggered when they are brought into large molecular structures by several receptors on the cell surface including the T-cell receptor (TCR), toll-like receptors (TLRs) and the death receptors (DRs).48 RIPK1 and RIPK3 are essential components of necrosomes.49 The oligomerized mixed lineage kinase domain-like protein (MLKL) invades and permeates the cell membrane, resulting in the death of the cell.50 Furthermore, the activation of RIP3-dependent necroptosis is also initiated by the cytosolic DNA sensor, which is a DNA-dependent activator of interferon (DAI) regulatory factors.51 Necroptosis is a cell death process that exposes the necrotic characteristics including membrane rupture and the depletion of organelles. Necroptosis can be determined by evaluating the disruption of the plasma membrane using dyes that cannot enter the cell, the discharge of cellular components such as cyclophilin A, high mobility group box 1 (HMGB1) and lactate dehydrogenase (LDH) through Western blot analysis, the measurement of mitochondrial potential using fluorescent probes and the investigation of cellular morphology using electron microscopy. Several alternative approaches including blockers that inhibit necroptosis and measurement of key proteins in the pathway can be used.52
Nevertheless, a newly discovered GSDMD protein has been uncovered and thoroughly investigated. The protein is commonly observed in a condition of autoinhibition.53 After caspase cleavage, GSDMD generates the N-terminal fragment (GSDMD-NT), which causes cells to swell and burst. Therefore, gasdermin D (GSDMD) acts as an effector molecule for pyroptosis-induced cell death. Comparable to the GSDMD pore-forming protein, DFNB59, DFNA5/GSDME, GSDMC, GSDMB and GSDMA activate pyroptosis and induce the denaturation of cytoplasmic membrane.54,55 Wang et al showed pyroptosis is triggered by GSDMD-NT and correlates with the interaction between 4 5-diphosphate phosphatidylinositol and the N-terminal domain of GSDME. This interaction leads to the creation of pores in liposomes and the removal of their phospholipid components.37 The study by Shi et al renamed pyroptosis as a gasdermin family-dependent programmed necrosis.40 Recently, Chauhan et al discovered that neutrophil elastase (NE) split GSDMD and caused neutrophil pyroptosis.56 Pyroptosis is a type of regulated cell death (RCD) that relies heavily on the gasdermin protein family to form pores in the cell membrane. This process is usually triggered by the activation of inflammatory caspases, as stated by the Nomenclature Committee on Cell Death (NCCD) in 2018.57
Molecular Mechanisms of Pyroptosis
Pyroptosis is induced by classic and non-classical inflammasome pathways, caspase-dependent apoptosis and granzymes-dependent signaling pathways.43 Gasdermin proteins are essential mediators in various cellular signaling pathways and must undergo cleavage by granzymes or precursor caspases.58 Caspases are classified into apoptotic and inflammatory caspases based on their distinct functions.59 Caspases-1/4/5/11 are inflammatory caspases that further suppress pathogen proliferation, regulate pyroptosis and release inflammatory substances.60 Caspases are primarily involved in protecting the body against infectious infections. Inflammasome is a multiprotein complex, which further activates the caspase-1 pathway. Caspase-1 is a member of the inflammatory caspase family, responsible for converting IL-18 into its fully developed state, which is a potent pro-inflammatory mediator. These pro-inflammatory variables distinguish pyroptosis from apoptosis, even though both processes rely on caspases.
Therefore, pyroptosis is lytic and characterized by cell ruptures, swellings and many bubbles blowing on the plasma membrane.61 Apoptotic caspases mainly induce and regulate the biological processes involved in programmed cell death, known as apoptosis. The cleavage of gasdermins by proteases leads to the activation of the pyroptosis-regulated cell death.62
Classical Signaling Pathways (Caspase-1-Dependent Pyroptosis)
The stimulation of inflammasomes may initiate classical pyroptotic cell death, which results in the fragmentation of GSDMD and the release of several pro-inflammatory molecules such as IL-18 and IL-1β.63,64 Inflammasomes are intricate assemblies of several molecules that activate the adaptive immune response and act as a defense mechanism against microbial infections.65–69 Inflammasomes can trigger illnesses that are not caused by microorganisms.70,71 Accumulating evidence indicates that inflammasomes and their associated cytokines play an important role in the progression of cancer including processes such as cell proliferation, invasion and metastasis.72–75 Cytosolic pattern recognition receptors (PRRs) that have been activated identify molecular patterns associated with pathogens and danger (DAMPs and PAMPs) to develop the inflammasome.76,77 Activation ofPRRs leads to the initiation of subsequent signaling pathways, resulting in the synthesis of type I interferons and several pro-inflammatory cytokines. PRRs bind to pro-caspase-1 and apoptosis-associated speck-like protein (ASC) to generate inflammasomes in response to cellular activation via signal molecules such as viruses and bacteria.78–80 The most prevalent PRRs are nucleotide-binding oligomerization domain-like receptors (NLRs), which include NLRP1, NLRP3 and NLRC4, missing in melanoma 2 (AIM2) and pyrin.81,82 The N-terminal pyrin domain (PYD), nucleotide-binding oligomerization domain (NOD), leucine-rich repeat (LRR) and caspase recruitment domain (CARD) are constituents of NLRP1.83 The PYD is crucial for engaging with the ASC protein. NOD stimulates the signal by controlling the production of ATP. LRR recognizes and automatically inhibits ligands. The CARD proteins then recruit pro-caspase-1. The presence of the anthrax lethal toxin, muramyl dipeptide and Toxoplasma gondii components might potentially stimulate the activation of NLRP1.84 NLRP3 consists of N-terminal PYD, NOD and LRR domains but does not have a CRAD domain. The NLRP3 inflammasome signaling axis is activated by several stimuli comprising fungi, viruses, bacteria, ATP, ROS, uric acid and intrinsic damage signals.85,86 The stimulation of the P2X7 receptor by extracellular ATP leads to the generation of IL-1β and the activation of the caspase-1 pathway. Thus, the outflow of K+ ions is enhanced.87 The NLRC4 protein consists of three domains: an N-terminal caspase activation and recruitment domain (CARD), a central nucleotide-binding domain (NBD) and a C-terminal leucine-rich repeat (LRR) domain. NLRC4 is upregulated by flagellin and proteins from type III secretion systems.88 The PYD and HIN-200 domains present in AIM2 can identify double-stranded nucleotides that originate from bacteria or viruses.89 The Pyrin protein consists of a PYD domain, two B-box domains and a C-terminal SPRY/PRY region. Pyrin specifically designates the toxins of bacteria or effectors that deactivate host Rho guanosine triphosphatases.90 PRRs either directly or passively attract pro-caspase-1 together with ASC to create the caspase-1-dependent inflammasome. This inflammasome then undergoes self-cleavage to activate caspase-1. The activation of the caspase-1 pathway results in the formation of the GSDMD-NT protein, which forms pores in the cytoplasm. Inflammation and pyroptosis are induced by this process (Figure 1).79 Inflammasome-mediated pyroptosis significantly protects immune cells from microbial infections.
 |
Figure 1 Cellular and molecular mechanisms of pyroptosis-dependent signaling pathways. Pyroptotic signaling pathways are mainly triggered by the stimulation of damage-associated molecular patterns (DAMPs) and pathogen-associated molecular patterns (PAMPs), leading to the activation of a variety of inflammasome components. The activated inflammasome proteins further activate the caspase-1 pathway. Then, the activated caspase-1 splits GSDMD to produce GSDMD N-fragment and plasma membrane pores, resulting in pyroptosis-dependent cell death. Furthermore, the caspase-1 pathway triggers the formation and release of IL-1β and IL-18 inflammatory factors. In addition, LPS binds to caspase-4/5/11 precursor, inducing pyroptosis. Caspase-3/GSDME can also cause pyroptosis-mediated cell death. Mitochondrial and death receptors can also trigger the caspase-3 pathway. The activated caspase-3 splits GSDME to produce GSDME N-fragment, creating plasma membrane pores, cell contraction and rupture and resulting in pyroptosis-mediated cell death. |
Non-Classical Signaling Pathways (Caspase-4/5/11-Dependent Pyroptosis)
The non-classical pyroptotic signaling pathway does not include the association of human caspase-4/5 (or its mouse equivalent, caspase-11) with the downstream sensory complexes. Human caspase-4/5 (equivalent to mouse caspase-11) may be activated by straightaway binding to intracellular LPS via the N-terminal CARD in the non-classical pyroptotic route, which does not include upstream sensory complexes.91 Unlike dendritic cells, macrophages are responsive to the oxidized phospholipid 1-palmitoyl-2-arachidonoyl-sn-glycero-3-phosphorylcholine (oxPAPC), which acts as a TLR4 agonist and suppresses the non-classical inflammasome.92 Caspase-4/5/11 may cleave GSDMD, resulting in the formation of GSDMD-NT. GSDMD-NT undergoes polymerization and forms cytoplasmic pores.93,94 The NLRP3/caspase-1 pathway is necessary for maturing and releasing IL-1β/IL-18, but caspase-4/5/11 cannot mature pro-IL-1/pro-IL-18.95,96 Furthermore, caspase-4/5/11 cleaves GSDMD, inducing the release of K+ ions and initiating the activation of NLRP3 inflammasomes and pyroptosis in the cells.97,98 Yang et al discovered that pannexin-1 is a crucial protein that triggers caspase-11-dependent non-classical pyroptotic cell death.97 Lipopolysaccharide (LPS) triggers the activation of caspase-11, which leads to the cleavage and modification of Pannexin-1. This process further leads to the extensive release of cellular ATP and the subsequent activation of pyroptosis via activating the purinergic (P2X7) receptor.99 In 2011, it was revealed that murine BMDMs lacking Pannexin-1 may cause the release of potassium ions and activate caspase-1 via the NLRP3 inflammasomewithout relying on P2X7.100 In addition, the elimination of pannexin-1 in mice protects to counteract endotoxin shock, suggesting that particular potassium (K+) channels control the non-classical NLRP3 inflammasome signaling pathway.97 Therefore, the stimulation of caspase-11 activates the NLRP3 inflammasome through non-classical pyroptotic signaling.101
Alternative Signaling Pathways (Caspase-3/8-Dependent Pyroptosis)
The composition of gasdermin proteins is highly conserved among members of the family. Gasdermins, except DFNB59, possess both C-terminal and N-terminal domains. Activation of the N-terminus results in pyroptosis.58 Previous studies have shown that chemotherapeutic drugs may trigger the activation of caspase-3, leading to the cleavage of the gasdermin E (GSDME) protein. This process causes an increase in GSDME expression and results in the formation of N-GSDME termini in cancer cells.37,102 Caspases involved in apoptosis Yersinia infection in mouse macrophages have been shown to hinder the activity of TGF-β-activated kinase 1 (TAK1) and initiate the cleavage of GSDMD via the caspase-8 signaling pathway.103,104 It was believed that caspases-3/8 could not generate gasdermin to induce pyroptosis. Further investigation revealed that Yersinia infection leads to the production of yersinia outer protein J (YopJ), a protein that hinders TAK1 and initiates the cleavage of GSDMD via caspase-8 in mouse macrophages.105 Therefore, these results enhance the advancement and broadening of information about pyroptosis. Unexpectedly, Hou et al revealed that programmed cell death ligand (PD-L1) regulates the process of converting TNF-induced apoptosis to pyroptosis in breast cancer cells.106 During hypoxia, the activation of p-Stat3 increases the movement of PD-L1 into the nucleus and the production of gasdermin C (GSDMC). LPS induces pyroptosis by the activation of caspase-4/5/11 signaling pathways.107 Scientists have explored that LPS stimulates macrophages to initiate pyroptosis by activating the caspase-8 pathway.106 The stimulation of TNF-α induces caspase-8 to lyse GSDMC, generate N-GSDMC and form membrane pores, resulting in pyroptosis.106 TNF induces pyroptosis in macrophages by activating nuclear PD-L1, caspase-8 and GSDMC. Furthermore, Hou et al showed that chemotherapeutics and antibiotics can enhance pyroptotic cell death of BC cells by activating caspase-8/GSDMC-dependent pyroptosis.106 Caspase-6 induces the NLRP3/caspase-1-dependent pyroptotic signaling axis by facilitating the connection among Z-DNA protein 1 and receptor-interacting serine/threonine protein kinase 3.108 Further investigation is highly required to determine the precise roles and activities of the additional caspases implicated in pyroptosis.
Researchers in 2020 discovered that chimeric antigen receptor (CAR) T cells secrete granzyme B (GzmB), which activates caspase-3.109 The activation of caspases-3 and GSDME pathways induces pyroptosis. Studies have shown that GzmB directly cleaves GSDME and triggers pyroptosis, which promotes the anti-tumor immune response and inhibits the growth of tumors.110 Cytotoxic T lymphocytes (CTLs) and natural killer (NK) cells were found to eliminate GSDMB-positive cells by triggering the process of pyroptosis. A recent study by Zhong et al reports that GSDMB is highly expressed in specific tissues, particularly inside the gastrointestinal system epithelium and tumors originated in this region.111 Zhou et al showed that gasdermin might go through hydrolysis at a site besides aspartic acid, leading to the formation of cytoplasmic pores.112 These results contradict the widely accepted belief that caspases may only trigger pyroptosis.
Role of Pyroptosis-Regulated Signaling Pathways in the Pathogenesis and Progression of EC
Inflammasomes are important components of the innate immune response to PAMPs and DAMPs.65 The NLRP3 inflammasome can react to a broad spectrum of stimuli. NLPR3 binds pro-caspase-1 through the pyrin domain of the adaptor protein of ASC, which promotes pro-IL-1β and pro-IL-18 to accelerate their maturation. Recent studies suggest that the NLRP3 inflammasome plays a significant role in the progression of several types of cancer including gastric cancer, breast cancer, colorectal cancer and liver cancer.74,113–115 Therefore, the prognosis for patients with EC is enhanced by investigating alternative cell death mechanisms. Patients diagnosed with EC exhibit elevated levels of oxidative stress (OS) and systemic inflammatory conditions, which can be attributed to the presence of metabolic syndrome. The significant role of inflammatory pathways in estrogen metabolism in EC is noteworthy.116,117 Radiotherapy and chemotherapy may not be effective in the treatment of serous carcinomas and tumors with high-risk characteristics such as distant metastases or deep myometrial invasion, as they are resistant to necrosis and apoptosis.118 Therefore, targeting pyroptotic cancerous cell death might be considered an alternative therapeutic avenue for the treatment and management of EC in the near future. However, it was found that the protein expression level of GSDMD varies significantly between EC cells and normal endometrial tissues, according to the data obtained from the TCGA database. Furthermore, this discrepancy was shown to be associated with the pathological type, stage and patient weight. Contemporary studies indicate that pyroptosis is a distinguishing feature of autoinflammatory and autoimmune disorders.117,119 Yang et al found that EC lesions exhibit pyroptotic events, as indicated by the presence of increased NLRP3, caspase-1 and GSDMD concentrations.120 The GSDMD-NT portion facilitates the formation of cytoplasmic pores that function as “hydrogen channels” for hydrogen efflux. Chang et al discovered that patients with EC had elevated expression of inflammasomes such as AIM2 and NLRP3 compared to the control group.121 The protein expression levels of GSDMD, NLRP3, caspase-1 and IL-1β were markedly elevated in atypical hyperplasia and cancer tissues compared to benign endometrial tissues. Therefore, GSDMD may play a significant role in the progression of EC by regulating pyroptosis (Figure 2).
 |
Figure 2 The pathophysiological mechanisms of pyroptosis-related signaling pathways in the progression of EC. |
The precise biochemical pathways implicated in tumor-associated macrophages (TAMs) polarization in extracellular matrix cancer (EMC) are not fully defined. It is widely believed that IL-1β modulates the immune response against tumors and triggers an inflammatory response.122,123 Immune cells such as lymphocytes, antigen-presenting cells, macrophages and neutrophils release several pro-inflammatory mediators such as IL-1β and IL-18.123 The activation of IL-1β is regulated by inflammasomes such as NLRP3, which further facilitate the conversion of pro-IL-1β into its active form.124 Abais et al indicated that the activation of OS can activate the NLRP3 inflammasome axis, resulting in the overproduction of ROS, specifically in macrophages.125
The identification of safe and effective treatments for activating pyroptosis in EC is of paramount importance. ROS is an upstream mechanism that mediates cellular pyroptosis.126 The production levels of ROS are drastically elevated in EC compared to normal endometrial tissue. Elevated ROS can further induce cell death by stimulating signaling pathways associated with pyroptosis in EC cells.127 The NLRP3 inflammasome induces damage to the host by inflammatory events including TNF-α exposure, NF-κB pathway activation and ROS accumulation.125 The expression of inflammasome components including NLRP3 and caspase-1 was significantly augmented in EC tumor tissues compared to benign tissues. ROS are bioproduct molecules characterized by their high reactivity, which can further induce OS and damage several physiological constituents.128 In addition, ROS activate intracellular signaling pathways, which promote cell proliferation, survival and migration.129 Accumulating evidence indicates that the excessive accumulation of ROS within cells can induce cell damage, cellular dysfunction and cell death.130–132 Several lines of studies have revealed that NLRP3 inflammasomes regulate ROS production through different regulatory mechanisms. This can either inhibit tumor growth by activating cellular stress and death or increase angiogenesis, inflammation and immune suppression, which can advance tumors.133–135 Recently, Zhu et al revealed that macrophage NLRP3 deletion reduced cytotoxic T cells in EM tumors, possibly due to altered TAM-lymphocyte interactions.136 The study confirms prior findings that macrophage NLRP3 signaling regulates the differentiation of T cells.
The immune-regulating thioredoxin interacting protein (TXNIP) has been associated with cardiovascular and neurological disorders.137–140 Studies have consistently indicated that TXNIP is downregulated in a variety of cancers.141 Several lines of studies also suggest that TXNIP expression is decreased in tumors.142–144 An innovative investigation carried out by Kim et al revealed that downregulation of TXNIP may further accelerate the progression of EC.145 Therefore, the TXNIP gene is considered a potential tumor suppressor.146 Growing evidence indicates that the expression of TXNIP is markedly downregulated in several cancers including lung, liver and breast cancers.147–149 The upregulation of TXNIP has been demonstrated to hinder the proliferation of cancerous cells.150 TXNIP has profound pathogenic significance and is interconnected with the cellular response to ROS. Normal cells typically produce low and stable levels of ROS, which are naturally occurring byproducts of oxygen metabolism. Therefore, the ROS/TXNIP signaling axis may play a contributory role in the progression of EC.
Several mechanisms have been implicated in the loss of viability of cells including autophagy and pyroptosis.151 Multiple pathological mechanisms are involved in the activation of pyroptosis.152 Researchers have discovered that pyroptosis can be used as a biomarker for diagnostic and predictive purposes in uterine corpus endometrial carcinoma (UCEC).153,154 Previous studies have linked pyroptosis to the advancement of cancer.116 Yu et al found a correlation between the glutamyl hydrolase (GGH) expression level of T helper type 2 (Th2) cells and the low infiltration of cell adhesion molecule 56 (CD56) killer cells.155 This association suggests a mechanism for driving the progression of UCEC. Zheng and his colleagues report that neural PAS domain protein 2 (NPAS2) can further induce UCEC by infiltrating immune cells into tumors.156 Liu et al designed and validated a Treg-related risk signature (TRRS) for predicting the outcome of UCEC and immune status.157 TRRS could be used to provide personalized treatment for UCEC patients based on their prognosis. A comprehensive review of the references revealed that pyroptosis is a significant factor in the advancement of inflammatory or malignant tumors. Intriguingly, Huang et al identified 47 genes that contribute to the progression of UCEC by regulating pyroptosis-related signaling pathways.154 Furthermore, 42 genes associated with pyroptosis were validated for their essential role in the development and progression of UCEC. Eight pyroptosis-related genes were used to develop pyroptosis-mediated gene models to predict the survival of patients with UCEC.
Research has shown that interferon regulatory factor 2 (IRF2) promotes the stimulation of GSDMD by inducing pyroptosis.158 Inflammatory vesicles activate the caspase-1 pathway, which further leads to focal death. It is believed that pyroptosis is caused by caspase-11 substrates that generate non-selective pores in the cytoplasmic membrane region, resulting in cell enlargement, ruptures and the release of proinflammatory mediators.25,159,160 A growing body of evidence indicates that GSDMD and GSDMB play a key role in the activation of pyroptosis in cancer, which enhances the preventative potential of UCEC.161,162 Selenoprotein GPX4 is a glutathione peroxidase with inhibitory properties for lipid peroxidation.163 It has been proven that GPX4 activates cancerous cells to initiate apoptosis.164–166 Guerriero et al demonstrated that GPX4 suppressed macrophage pyroptosis in mice model.167 The augmented level of GPX4 could efficaciously suppress pyroptosis in UCEC. Targeting the GPX4 inducers may alleviate the symptoms of UCEC associated with pyroptosis and improve patient survival. Previous studies have demonstrated that charged multivesicular body protein 2A (CHMP2A) deficiencies can promote the formation of autophagic vesicles and activate pyroptosis.168 The actual role and function of CHMP2A in EC remains unclear. Current studies have revealed that the identification of low levels of CHMP2A expression in UCEC patients is associated with poor survival, suggesting that inhibiting CHMP2A could serve as a therapeutic target in this disease.154 Therefore, pyroptosis-related genes have a significant role in the pathogenesis and advancement of UCEC.
The innate immune response to bacterial LPS relies on toll-like receptor 4 (TLR4) and nuclear factor kappa B (NF-kB) pathways.169,170 TLR4 senses LPS and activates the NF-kB signaling pathway, which modulates the creation of several pro-inflammatory mediators and immune response genes through the NLRP3 inflammasome domain.171 In addition, macrophages activate the TLR4/NF-kB pathway via NLRP3 inflammasome, which may alter pro-inflammatory molecules and other immune-regulating genes.172 EMC cells co-cultured with NLRP3-depleted macrophages exhibit increased growth, invasion, and migration potential.136 It was observed that the expression level of NLRP3 was downregulated in EMC macrophages. NLRP3 depletion might be a key molecular mechanism responsible for the impact of cancer biology on macrophages.
Caspases 11 is one of the exciting members of the Caspases family. Nobuhiko et al confirmed that Caspases sense bacterial LPS, mediate immune responses and trigger pyroptosis.173 LPS and caspase-11 bind to lipid A in gram-negative bacteria with their caspase-activated recruitment domain (CARD).174 LPS and TLR4-MD2 display a similar affinity through electrostatic adsorption to most of the positive charges found in the CARD domain.175 Caspases-11 can recognize LPS directly without an NRL-like scaffold. Caspase-11 fragments when LPS interacts with its CARD domain.176 TLRs are activated in inflammatory tissues, which further induces cell death by activating caspase-11.177 The strong association between gram-negative anaerobes and periodontitis indicates that caspase-11-dependent pyroptosis plays a significant part in the advancement of EC. Therefore, caspase-11 has excellent potential as a focus for therapeutic intervention in the treatment and management of EC (Figure 3). Nevertheless, the actual function of caspase-11-dependent pyroptosis in the development of EC remains unknown. The significance of caspase-11 in EC needs to be further investigated and confirmed.
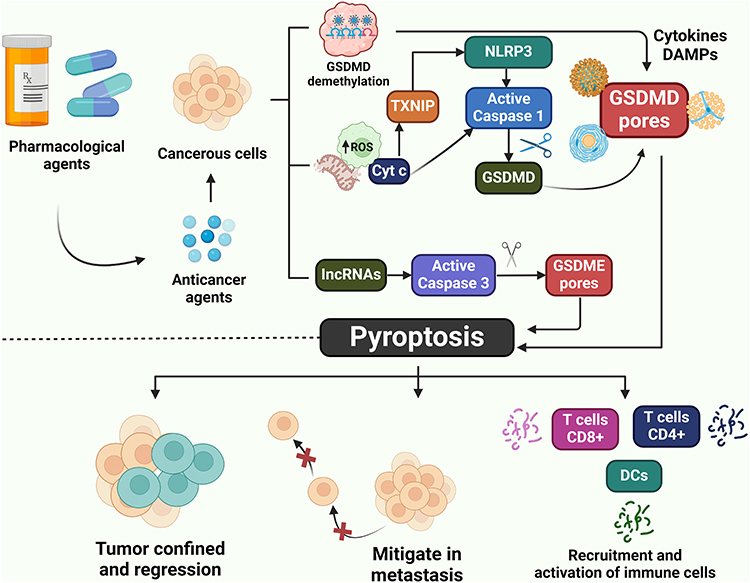 |
Figure 3 The schematic strategy for the pharmacological induction of pyroptosis-mediated cancer cell death for the therapeutic regulation of EC. |
Endogenous ATP binds to P2X7 receptors, which are present at high nanomolar and low micromolar levels in the extracellular fluid of epithelial cells.178 P2X7 is a receptor for P2 nucleotides located on the cell membrane and is activated by ligands. Extracellular ATP activates the P2X7 receptor, which creates channels in the plasma membrane that allow the passage of cations up to 900 kDa. Multiple evidence indicates that stimulation P2X7R facilitates the formation of NLRP3, which is an intracellular multimeric protein complex that triggers inflammatory responses and cell death (pyroptosis and apoptosis) by activating caspase-1.36,179–182 Caspase-1 pathway activation fosters GSDMD-mediated pyroptosis, leading to the production of pro-inflammatory mediators including IL-1β and IL-18.183 Li et al have shown that the expression of the P2X7 receptor is downregulated in epithelial cancer cells of the ectodermal, urinary sinus, and distal paramesonephric duct.184 Kim et al demonstrated that epithelial cancer cells from the ectodermal, urogenital sinus, and distal paramesonephric ducts express reduced P2X7 level.185 Therefore, the downregulation of P2X7-dependent pyroptosis contributes to the progression of EC.
The endometriosis syndrome is a chronic inflammatory condition associated with a mechanism reliant on estrogen that affects approximately 10% of pregnant women.186 Endometriosis of the ovary is the predominant manifestation of endometriosis and can induce sterility and dysmenorrhea.187 The inflammasome is a complex composed of several proteins including Nod-like receptors (NLRs). These proteins are responsible for detecting pathogen-associated molecular patterns and molecular processes that disrupt homeostasis. Inflammasomes such as NLRP3, which contain nucleotide-binding oligomerization domains, leucine-rich repeats and pyrin domains, have been implicated in the development of endometriosis.188–191 Recently, Zhang et al showed that NLRP3 inflammasome-mediated pyroptosis induces Notch signal activation in endometriosis.192 In addition, Hang et al have reported that E3 ubiquitin ligase tripartite motif-containing 24 protein (TRIM24) deficiency promotes NLRP3/caspase-1/IL-1β-mediated pyroptosis in endometriosis.193 Intriguingly, Zhao et al suggest that astrocyte elevated gene-1 (AEG-1) exacerbates inflammation by facilitating the development of NLRP3 inflammasome in endometrial lesions in mice.194 The NLRP3 receptor is an intracellular receptor that detects both external and internal stimuli. The NLRP3 inflammasome complex is formed by the interaction of this substance with ASC and caspase-1. The combination stimulates the secretion of pro-inflammatory cytokines IL-1β and IL-18, enhancing the immune response and pyroptosis.35 Prior research has shown the distinct function of NLRP3 inflammasome in triggering the activation of mast cells (MCs) during the autoinflammatory response.195 Anti-cytokine therapy targets NLRP3 inflammasome activation and IL-1β production.196 Nevertheless, the precise processes by which the NLRP3 inflammasome influences the formation of ectopic endometrium remain unclear. The groundbreaking research of Guo et al has shown that endometriosis is associated with the activation of the NLRP3 inflammasome by estrogen via a nuclear-initiated signaling pathway.197 In summary, inflammasome and pyroptosis contribute to the development and advancement of endometriosis.
Molecular and genomic profiling in EC is becoming increasingly popular.198–200 The L1 cell adhesion molecule (L1CAM) is frequently mutated in endometrial cancer.201 Kommoss et al have demonstrated L1CAM) to be a significant indicator of high-risk disease in EC.202 Recently, Giannini et al reported that L1CAM has a prognostic role in stage EC, thus providing a potentially useful tool for tailoring the need for adjuvant therapy.203 L1CAM expression influences survival outcomes in stage-I EC. Recently, it has been shown that specific genetic markers L1CAM, Annexin 2, insulin-like growth factor receptor, epidermal growth factor receptor, etc) and aberrant molecular signaling pathways could be key players in metastatic processes in EC cells, although further clinical trials are required to confirm their prognostic value of EC in clinical practice.204,205 It was found that high L1CAM expression correlated with worse disease-free survival (HR 4.11, 95% CI 1.02–16.59, p = 0.047) and overall survival (HR 3.62, 95% CI 1.32–9.31, p = 0.012). High L1CAM level was also associated with a more aggressive FIGO grade and with older age. However, the role and significant relationship of L1CAM with pyroptosis is still unclear. The published paper indicates that L1CAM-regulated inflammasome or pyroptosis may play a possible role in EC progression. However, further studies are highly required to elucidate the contributory role of L1CAM-regulated pyroptosis in the pathogenesis and progression of EC.
Therapeutic Regulation of EC via Pyroptosis-Dependent Signaling Pathways
Hydrogen (H2) may suppress tumor formation and protect normal healthy cells. Recent studies have shown that H2 can be applied in the treatment of a variety of diseases including cancer, metabolic diseases and organ ischemia/reperfusion injury.206–208 Numerous studies also suggest that H2 impedes tumor cell activity, proliferation, invasion and migration dose- and time-dependently and promotes apoptosis in cervical, breast and cutaneous melanoma.209–211 Further evidence also indicates that H2 can significantly alleviate the progression of lung cancer, colon cancer, ovarian cancer, thymic lymphoma, Ehrlich ascites tumor cells, oral squamous cell carcinoma (OSCC) and fibrosarcoma cells, reduce tumor volume and weight and suppress tumor growth in xenografted mice.209,212–215 In addition, H2 has been shown to mitigate radiotherapy and chemotherapy-induced renal toxicity.216 The published work of Liu et al showed that H2 stimulates pyroptosis by activating the ROS-NLRP3-caspase-1 signaling pathways in the EC.120 Thus, H2 induces GSDMD-dependent pyroptosis, which may enhance the effectiveness of cancer therapies that target GSDMD. The pharmacological efficacy of H2 in inducing pyroptosis-dependent cancer cell death for therapeutic regulation of EC requires further study.
Inflammasomes consist of NLR family proteins, leucine-rich repeat and pyrin domain-containing (NLRP) 1b, NLRP9b, NLRP6, NLRP3, or NLR family caspase recruitment domain (CARD)-containing protein (NLRC) 4, which sense danger-related, pathogen-related and homeostasis-altering molecular patterns.217,218 Prior research has uncovered that the NLRP3 inflammasome contributes to the development of endometriosis.219 Several lines of studies have indicated that prolonged endometriosis can further result in the advancement of EC.220,221 The NLR NLRP7 is associated with myometrial invasion in human endometrial cancerous tissues and with endometriosis. Furthermore, endometrial cyst contents stimulate NLRP3 inflammasome in a stressful environment more strongly than cultured cells. The pharmacological strategy of suppressing the overactivation of NLRP3 inflammasome may be more effective in the management of EC. MCC950 can not hinder the major anti-microbial inflammasome components including NLRC4 and NLRP1. Osoku et al showed that the expression level of NLRP3 was significantly augmented compared to other NLRs in OE samples and CSCs.121 Researchers have discovered that overactivation of the inflammasome and proteins involved in pyroptosis leads to cell death, alleviates the development of EC and offers potential therapeutic targets. Furthermore, the researchers discovered that MCC950 effectively suppressed the activation of oxidative stress in granulosa cells of the mouse model with endometriosis. However, the exact pharmacological effects and underlying molecular mechanisms of NLRP3 inflammasome blocker MCC950 in alleviating the progression of EC remains unclear. Therefore, extensive research is strongly required to explore the pharmacological actions of MCC950 to discover promising anti-cancer drugs for the treatment and management of EC.
Long non-coding RNA (lncRNA) refers to RNA fragments longer than 200 nucleotides that are not genetically coded for protein synthesis.222,223 The significant role and function of lncRNA in biology was previously thought to be undefined. It has recently been demonstrated that lncRNA plays an important role in the regulation of cancer. Current studies indicate that lncRNA may also serve as a biomarker for the detection and prediction of a variety of cancers including EC.224,225 Studies have revealed that the long non-coding RNA small nucleolar RNA host gene 4 (SNHG4) regulates the epithelial-mesenchymal transition (EMT) signal and exhibits antitumor properties in esophageal cancer by acting on specificity protein 1 (SP-1) transcription factor.226 It has been demonstrated that long non-coding RNA SLERT controls the BDNF/TRKB pathway to facilitate the spread of EC cells.227 Further research into lncRNAs will provide new insight into the potential role of new lncRNAs in EC. Recently, Shan et al reported that ENST00000534735 suppresses the proliferation and migration of EC cells and promotes programmed cell death and inflammatory cell death by the OSBPL3 protein via regulating the APMK/SIRT1/NF-κB signaling pathway.228 It was found that the high ENST00000534735 expression resulted in a considerable elevation in levels of cleaved caspase-1, GSDME-N and NLRP3. Conversely, the reduction of ENST00000534735 led to a decrease in the expression of cleaved caspase-1, GSDME-N and NLRP3 in HEC-1A and Ishikawa cells.
Previously, Mao et al revealed that the TFEB-ERRα pathway stimulates the alteration of lipid composition and promotes the development of pseudopodia in EC cells, resulting in enhanced cell membrane fluidity. This process facilitates the invasion and metastasis of EC.229 Further studies are required to fully understand the process by which ERRα promotes resistance to chemotherapy. Tumor cells have an enhanced glycolytic capacity in order to meet their higher energy requirements. Furthermore, there is a correlation between the rate of aerobic glycolysis and the development, advancement and resistance to drugs in cancer.230 Zeng et al found that miR-211-5p promotes cancerous effects in low-metastatic melanoma tumor cells by inhibiting pyroptosis and increasing glycolysis. This is achieved by modulating the expression of the targeted gene guanine nucleotide-binding protein subunit α-15 (GNA15).231 ERRα plays an important role in regulating energy metabolism. Analysis of the cancer genome atlas (TCGA) dataset revealed that genes associated with ERRα are engaged in the process of glucose metabolism and regulated cell death. The data clearly suggested that the excessive production of ERRα may stimulate glycolysis, elevate the ECAR in EC cells and promote pyroptotic resistance. This may indicate the primary interaction between the metabolic shift towards glycolysis and the activation of pyroptosis in endothelial cells triggered by chemotherapy. ERR+ cells exhibited a high IC50 for cisplatin (DDP) compared to EC cells lacking ERR+. It has been shown that the overexpression of ERRα causes resistance to DDP by suppressing pyroptosis and inducing glycolytic reprogramming in EC cells. It was discovered by Daniela et al that ovarian cancer cells that are resistant to chemotherapy showed increased sensitivity to glucose deprivation. The drug-resistant cells also rely heavily on glucose to maintain their viability.232 A study revealed that hepatocellular carcinoma cells exhibited an increase in glycolytic metabolism, which is associated with resistance to sorafenib.233 He and his colleagues examined the possible involvement of metabolic reprogramming in the development of drug resistance in osteosarcoma and the specific molecular mechanisms that regulate this phenomenon. The scientists demonstrated a correlation between elevated ERRα expression and the metabolic reprogramming of osteosarcoma cells that are resistant to treatment. Targeted suppression of ERRα expression reverses the metabolic mode transformation.234 Thus, the overexpression of ERR in patient-derived EC increases resistance to pyroptosis and upregulates glycolysis-related genes in tumor cells.
Conclusion, Limitations and Future Directions
Pyroptosis induced by caspases involves cytoplasmic pore formation, cell lysis, membrane denaturation and secretion of several intracellular components. Multiple diseases have been associated with pyroptotic cell death, which has attracted considerable attraction from researchers and clinicians. Recent studies have increasingly concentrated on the contributing roles and functions of inflammasome and pyroptosis-regulated cell death in EC tumors. Our study provided an understanding of the intricate molecular processes of pyroptosis-related signaling pathways, which may offer new therapeutic approaches for the treatment and management of EC. Thus, targeting pyroptosis can deliver an innovative strategy for the treatment and regulation of a variety of cancers including EC. Current researches suggest that pharmacologically inducing pyroptosis-dependent cancer cell death can efficaciously suppress the formation of malignant tumors and provide a new treatment option for EC. Currently, research focuses primarily on compounds that activate inflammasomes such as NLRP3 and caspase-1 and promote pyroptosis, which could be useful for the treatment of EC. There are still many unresolved issues in the study of pyroptosis. The present and future implications of pyroptosis-mediated cell death in EC research are only beginning to be explored. A limited body of research has specifically investigated the molecular regulator of GSDMD, a key pyroptosis executor in EC. Furthermore, the precious function and role of non-canonical inflammasome and GSDME-dependent pyroptotic signaling pathways in the development of EC have not yet been explored. Therefore, more research is highly required to understand and clarify the contributory role and function of molecular mechanisms of non-canonical inflammasome and apoptotic-mediated pyroptosis in the progression of EC. In addition, extensive research on pyroptosis should also be conducted to uncover new possibilities and avenues for the therapeutic regulation of EC. Specific potential candidates have shown promise as prospective therapeutic agents for the induction of pyroptosis-mediated cancerous cell death in the therapy and control of EC. However, additional investigation is required to comprehend the processes better and develop targeted therapies for inflammasome activation and pyroptosis in EC.
Funding
This review was partially supported by the Key Research and Development Program of Zhejiang Province (2024C03171), the Key Research and Development Project of Lishui (2023zdyf15), the Public Welfare Technology Research Funding Project of Zhejiang (LTGY24H100002 & LLSSY24H160003), the Zhejiang Provincial Medical and Health Science and Technology Project (2022RC302) and the Post-Doctoral Research Start-Up Fund of Lishui People’s Hospital, Lishui, Zhejiang, China (2023bsh001).
Disclosure
The authors have declared no competing interests.
References
1. Siegel RA-O, Miller KA-O, Jemal AA-O. Cancer statistics, 2019. CA: A Cancer Journal for Clinicians. 2019;69(1):7–34. doi:10.3322/caac.21551
2. Donnez J, Dolmans MM. Uterine fibroid management: from the present to the future. Human Reproduction Update. 2016;22(6):665–686. (). doi:10.1093/humupd/dmw023
3. Bellovin DI, Das B, Felsher DW, Felsher DW. Tumor dormancy, oncogene addiction, cellular senescence, and self-renewal programs. Systems Biology of Tumor Dormancy. 2013;2013:91–107
4. Shi J, Zhao Y, Wang K, et al. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature. 2015;526(7575):660–665. doi:10.1038/nature15514
5. Jorgensen I, Rayamajhi M, Miao EA. Programmed cell death as a defence against infection. Nature Reviews Immunology. 2017;17(3):151–164. (). doi:10.1038/nri.2016.147
6. Zeng C, Wang R, Tan H. Role of Pyroptosis in Cardiovascular Diseases and its Therapeutic Implications. Int J Bio Sci. 2019;15(7):1345–1357. doi:10.7150/ijbs.33568
7. Chen X, Tian PC, Wang K, Wang M, Wang K. Pyroptosis: role and Mechanisms in Cardiovascular Disease. Front Cardiovasc Med. 2022;9:897815. doi:10.3389/fcvm.2022.897815
8. Wang M, Wang K, Gao X, Zhao K, Chen H, Xu M. Anti-inflammatory effects of isoalantolactone on LPS-stimulated BV2 microglia cells through activating GSK-3β-Nrf2 signaling pathway. Int Immunopharmacol. 2018;65:323–327. doi:10.1016/j.intimp.2018.10.008
9. Jia C, Chen H, Zhang J, et al. Role of pyroptosis in cardiovascular diseases. Int Immunopharmacol. 2019;67:311–318. doi:10.1016/j.intimp.2018.12.028
10. Carneiro BA, El-Deiry WS. Targeting apoptosis in cancer therapy. Nature Reviews. Clinical Oncology. 2020;17(7):395–417. doi:10.1038/s41571-020-0341-y
11. Jan R, Chaudhry GE. Understanding Apoptosis and Apoptotic Pathways Targeted Cancer Therapeutics. Advanced Pharmaceutical Bulletin. 2019;9(2):205–218. doi:10.15171/apb.2019.024
12. Mortezaee K, Najafi M, Farhood B, et al. Modulation of apoptosis by melatonin for improving cancer treatment efficiency: an updated review. Life Sci. 2019;228:228–241. doi:10.1016/j.lfs.2019.05.009
13. Oladapo A, Jackson T, Menolascino J, Periyasamy P. Role of pyroptosis in the pathogenesis of various neurological diseases. Brain Behav Immun. 2024;117:428–446. doi:10.1016/j.bbi.2024.02.001
14. Wu C, Chen H, Zhuang R, et al. Betulinic acid inhibits pyroptosis in spinal cord injury by augmenting autophagy via the AMPK-mTOR-TFEB signaling pathway. Int J Bio Sci. 2021;17(4):1138–1152. doi:10.7150/ijbs.57825
15. Al Mamun A, Wu Y, Monalisa I, et al. Role of pyroptosis in spinal cord injury and its therapeutic implications. J Adv Res. 2021;28:97–109. doi:10.1016/j.jare.2020.08.004
16. Elias EE, Lyons B, Muruve DA. Gasdermins and pyroptosis in the kidney. Nature Reviews. Nephrology. 2023;19(5):337–350. doi:10.1038/s41581-022-00662-0
17. Cao Z, Huang D, Tang C, et al. Pyroptosis in diabetes and diabetic nephropathy. Int j Clin Chem. 2022;531:188–196. doi:10.1016/j.cca.2022.04.011
18. Mamun AA, Wu Y, Nasrin F, et al. Role of Pyroptosis in Diabetes and Its Therapeutic Implications. J Inflamm Res. 2021;14:2187–2206. doi:10.2147/JIR.S291453
19. Vande Walle L, Lamkanfi M. Pyroptosis. Curr Biol. 2016;26(13):R568–r572. doi:10.1016/j.cub.2016.02.019
20. Zychlinsky A, Prevost MC, Sansonetti PJ. Shigella flexneri induces apoptosis in infected macrophages. Nature. 1992;358(6382):167–169. doi:10.1038/358167a0
21. Chen Y, Smith MR, Thirumalai K, Zychlinsky A. A bacterial invasin induces macrophage apoptosis by binding directly to ICE. EMBO J. 1996;15(15):3853–3860. doi:10.1002/j.1460-2075.1996.tb00759.x
22. Hersh D, Monack DM, Smith MR, Ghori N, Falkow S, Zychlinsky A. The Salmonella invasin SipB induces macrophage apoptosis by binding to caspase-1. Proc Natl Acad Sci USA. 1999;96(5):2396–2401. doi:10.1073/pnas.96.5.2396
23. Cookson BT, Brennan MA. Pro-inflammatory programmed cell death. Trends Microbiol. 2001;9(3):113–114. doi:10.1016/S0966-842X(00)01936-3
24. Boise LH, Collins CM. Salmonella-induced cell death: apoptosis, necrosis or programmed cell death? Trends in microbiology. Trends in Microbiology. 2001;9(2):64–67. doi:10.1016/s0966-842x(00)01937-5
25. Chen X, He WT, Hu L, et al. Pyroptosis is driven by non-selective gasdermin-D pore and its morphology is different from MLKL channel-mediated necroptosis. Cell Res. 2016;26(9):1007–1020. doi:10.1038/cr.2016.100
26. Weindel CG, Ellzey LM, Martinez EL, Watson RO, Patrick KL. Gasdermins gone wild: new roles for GSDMs in regulating cellular homeostasis. Trends Cell Biol. 2023;33(9):773–787. doi:10.1016/j.tcb.2023.02.007
27. Fink SL, Cookson BT. Caspase-1-dependent pore formation during pyroptosis leads to osmotic lysis of infected host macrophages. Cellular Microbiology. 2006;8(11):1812–1825. doi:10.1111/j.1462-5822.2006.00751.x
28. Brennan MA, Cookson BT. Salmonella induces macrophage death by caspase-1-dependent necrosis. Mol Microbiol. 2000;38(1):31–40. doi:10.1046/j.1365-2958.2000.02103.x
29. Wang Q, Imamura R, Motani K, Kushiyama H, Nagata S, Suda T. Pyroptotic cells externalize eat-me and release find-me signals and are efficiently engulfed by macrophages. Int Immunol. 2013;25(6):363–372. doi:10.1093/intimm/dxs161
30. de Vasconcelos NM, Lamkanfi M. Recent Insights on Inflammasomes, Gasdermin Pores, and Pyroptosis. Cold Spring Harbor Perspect Biol. 2020;12(5):a036392. doi:10.1101/cshperspect.a036392
31. Rudel T, Bokoch GM. Membrane and morphological changes in apoptotic cells regulated by caspase-mediated activation of PAK2. Science. 1997;276(5318):1571–1574. doi:10.1126/science.276.5318.1571
32. Poon IK, Lucas CD, Rossi AG, Ravichandran KS. Apoptotic cell clearance: basic biology and therapeutic potential. Nat Rev Immunol. 2014;14(3):166–180. doi:10.1038/nri3607
33. Tang R, Xu J, Zhang B, et al. Ferroptosis, necroptosis, and pyroptosis in anticancer immunity. Journal of Hematology & Oncology. 2020;13(1):110. doi:10.1186/s13045-020-00946-7
34. Bertheloot D, Latz E. Necroptosis, pyroptosis and apoptosis: an intricate game of cell death. Cell Mol Immunol. 2021;18(5):1106–1121.
35. Swanson KV, Deng M. The NLRP3 inflammasome: molecular activation and regulation to therapeutics. Nat Rev Immunol. 2019;19(8):477–489.
36. He Y, Hara H, Núñez G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem Sci. 2016;41(12):1012–1021. doi:10.1016/j.tibs.2016.09.002
37. Wang Y, Gao W, Shi X, et al. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature. 2017;547(7661):99–103. doi:10.1038/nature22393
38. Kesavardhana S, Malireddi RKS, Kanneganti TD. Caspases in Cell Death, Inflammation, and Pyroptosis. Ann Rev Immunol. 2020;38(1):567–595. doi:10.1146/annurev-immunol-073119-095439
39. Liu X, Zhang Z, Ruan J, et al. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature. 2016;535(7610):153–158. doi:10.1038/nature18629
40. Shi J, Gao W, Shao F. Pyroptosis: gasdermin-Mediated Programmed Necrotic Cell Death. Trends Biochem Sci. 2017;42(4):245–254. doi:10.1016/j.tibs.2016.10.004
41. Purnama CA, Meiliana A, Barliana MI, Lestari K. Update of cellular responses to the efferocytosis of necroptosis and pyroptosis. Cell Divis. 2023;18(1):5. doi:10.1186/s13008-023-00087-6
42. Miao EA, Rajan JV, Aderem A. Caspase-1-induced pyroptotic cell death. Immunol Rev. 2011;243(1):206–214. doi:10.1111/j.1600-065X.2011.01044.x
43. Bergsbaken T, Fink SL, Cookson BT. Pyroptosis: host cell death and inflammation. Nat Rev Microbiol. 2009;7(2):99–109. doi:10.1038/nrmicro2070
44. Xie Y, Hou W, Song X, et al. Ferroptosis: process and function. Cell Death Differ. 2016;23(3):369–379. doi:10.1038/cdd.2015.158
45. Lewerenz J, Hewett SJ, Huang Y, et al. The cystine/glutamate antiporter system x(c)(-) in health and disease: from molecular mechanisms to novel therapeutic opportunities. Antioxid Redox Signal. 2013;18(5):522–555. doi:10.1089/ars.2011.4391
46. Conrad M, Kagan VE, Bayir H, et al. Regulation of lipid peroxidation and ferroptosis in diverse species. Genes & Development. 2018;32(9–10):602–619. doi:10.1101/gad.314674.118
47. Vanlangenakker N, Vanden Berghe T, Vandenabeele P. Many stimuli pull the necrotic trigger, an overview. Cell Death Differ. 2012;19(1):75–86.
48. He S, Liang Y, Shao F, Wang X. Toll-like receptors activate programmed necrosis in macrophages through a receptor-interacting kinase-3-mediated pathway. Proc Natl Acad Sci USA. 2011;108(50):20054–20059. doi:10.1073/pnas.1116302108
49. Wegner KW, Saleh D, Degterev A. Complex Pathologic Roles of RIPK1 and RIPK3: moving Beyond Necroptosis. Trends Pharmacol Sci. 2017;38(3):202–225. doi:10.1016/j.tips.2016.12.005
50. Cai Z, Jitkaew S, Zhao J, et al. Plasma membrane translocation of trimerized MLKL protein is required for TNF-induced necroptosis. Nat Cell Biol. 2014;16(1):55–65. doi:10.1038/ncb2883
51. Maelfait J, Liverpool L. Sensing of viral and endogenous RNA by ZBP1/DAI induces necroptosis. THE EMBO Journal. 2017;36(17):2529–2543. doi:10.15252/embj.201796476
52. Vanden Berghe T, Grootjans S, Goossens V, et al. Determination of apoptotic and necrotic cell death in vitro and in vivo. Methods. 2013;61(2):117–129. doi:10.1016/j.ymeth.2013.02.011
53. Devant P, Kagan JC. Molecular mechanisms of gasdermin D pore-forming activity. Nature Immunology. 2023;24(7):1064–1075. doi:10.1038/s41590-023-01526-w
54. Liu X, Xia S. Channelling inflammation: gasdermins in physiology and disease. Nature Reviews. Drug Discovery. 2021;20(5):384–405. doi:10.1038/s41573-021-00154-z
55. Ouyang X, Zhou J, Lin L, Zhang Z, Luo S, Hu D. Pyroptosis, inflammasome, and gasdermins in tumor immunity Innate Immun. 2023;29(1–2):3–13. doi:10.1177/17534259221143216
56. Chauhan D, Demon D, Vande Walle L, et al. GSDMD drives canonical inflammasome-induced neutrophil pyroptosis and is dispensable for NETosis. EMBO Reports. 2022;23(10):e54277. doi:10.15252/embr.202154277
57. Galluzzi L, Vitale I, Aaronson SA, et al. Molecular mechanisms of cell death: recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018;25(3):486–541. doi:10.1038/s41418-017-0012-4
58. Zou J, Zheng Y, Huang Y, Tang D, Kang R, Chen R. The Versatile Gasdermin Family: their Function and Roles in Diseases. Front Immunol. 2021;12:751533. doi:10.3389/fimmu.2021.751533
59. Van Opdenbosch N, Lamkanfi M. Caspases in Cell Death, Inflammation, and Disease. Immunity. 2019;50(6):1352–1364. doi:10.1016/j.immuni.2019.05.020
60. Man SM, Karki R, Kanneganti TD. Molecular mechanisms and functions of pyroptosis, inflammatory caspases and inflammasomes in infectious diseases. Immunological Reviews. 2017;277(1):61–75. doi:10.1111/imr.12534
61. Matikainen S, Nyman TA. Function and Regulation of Noncanonical Caspase-4/5/11 Inflammasome. Journal of Immunology (Baltimore, Md.: 1950). 2020;204(12):3063–3069. doi:10.4049/jimmunol.2000373
62. Heilig R, Dilucca M, Boucher D. Caspase-1 cleaves Bid to release mitochondrial SMAC and drive secondary necrosis in the absence of GSDMD. Life Science Alliance. 2020;3(6). doi:10.26508/lsa.202000735
63. He WT, Wan H, Hu L, et al. Gasdermin D is an executor of pyroptosis and required for interleukin-1β secretion. Cell Res. 2015;25(12):1285–1298. doi:10.1038/cr.2015.139
64. Schneider KS, Groß CJ, Dreier RF, et al. The Inflammasome Drives GSDMD-Independent Secondary Pyroptosis and IL-1 Release in the Absence of Caspase-1 Protease Activity. Cell Rep. 2017;21(13):3846–3859. doi:10.1016/j.celrep.2017.12.018
65. Rathinam VA, Fitzgerald KA. Inflammasome Complexes: emerging Mechanisms and Effector Functions. Cell. 2016;165(4):792–800. doi:10.1016/j.cell.2016.03.046
66. Lamkanfi M. Emerging inflammasome effector mechanisms. Nat Rev Immunol. 2011;11(3):213–220. doi:10.1038/nri2936
67. Jacobs SR, Damania B. NLRs, inflammasomes, and viral infection. J Leukocyte Biol. 2012;92(3):469–477. doi:10.1189/jlb.0312132
68. Zhang Y, Chi Z, Cui Z, Chang S, Wang Y, Zhao P. Inflammatory response triggered by avian hepatitis E virus in vivo and in vitro. Front Immunol. 2023;14:1161665. doi:10.3389/fimmu.2023.1161665
69. Tsuchiya K, Hara H. The inflammasome and its regulation. Critical Reviews in Immunology. 2014;34(1):41–80. doi:10.1615/CritRevImmunol.2013008686
70. Antushevich H. Interplays between inflammasomes and viruses, bacteria (pathogenic and probiotic), yeasts and parasites. Immunol Lett. 2020;228:1–14. doi:10.1016/j.imlet.2020.09.004
71. Zheng D, Liwinski T, Elinav E. Inflammasome activation and regulation: toward a better understanding of complex mechanisms. Cell Discovery. 2020;6:36.
72. Deng Z, Lu L, Li B, Shi X, Jin H, Hu W. The roles of inflammasomes in cancer. Front Immunol. 2023;14:1195572. doi:10.3389/fimmu.2023.1195572
73. Lillo S, Saleh M. Inflammasomes in Cancer Progression and Anti-Tumor Immunity. Front Cell Develop Biol. 2022;10:839041. doi:10.3389/fcell.2022.839041
74. Faria SS, Costantini S, de Lima VCC, et al. NLRP3 inflammasome-mediated cytokine production and pyroptosis cell death in breast cancer. Journal of Biomedical Science. 2021;28(1):26. doi:10.1186/s12929-021-00724-8
75. Jang J-H, Kim D-H, Surh Y-J. Dynamic roles of inflammasomes in inflammatory tumor microenvironment. Npj Precision Oncology. 2021;5(1). doi:10.1038/s41698-021-00154-7
76. Amarante-Mendes GP, Adjemian S, Branco LM, Zanetti LC, Weinlich R, Bortoluci KR. Pattern Recognition Receptors and the Host Cell Death Molecular Machinery. Front Immunol. 2018;9:2379. doi:10.3389/fimmu.2018.02379
77. Tang D, Kang R, Coyne CB, Zeh HJ, Lotze MT. PAMPs and DAMPs: signal 0s that spur autophagy and immunity. Immunol Rev. 2012;249(1):158–175. doi:10.1111/j.1600-065X.2012.01146.x
78. Jorgensen I, Zhang Y, Krantz BA. Pyroptosis triggers pore-induced intracellular traps (PITs) that capture bacteria and lead to their clearance by efferocytosis. The Journal of Experimental Medicine. 2016;213(10):2113–2128. doi:10.1084/jem.20151613
79. Miao EA, Leaf IA, Treuting PM, et al. Caspase-1-induced pyroptosis is an innate immune effector mechanism against intracellular bacteria. Nat Immunol. 2010;11(12):1136–1142. doi:10.1038/ni.1960
80. Hara H, Seregin SS, Yang D, et al. The NLRP6 Inflammasome Recognizes Lipoteichoic Acid and Regulates Gram-Positive Pathogen Infection. Cell. 2018;175(6):1651–1664.e1614. doi:10.1016/j.cell.2018.09.047
81. Wu J, Fernandes-Alnemri T, Alnemri ES. Involvement of the AIM2, NLRC4, and NLRP3 inflammasomes in caspase-1 activation by Listeria monocytogenes. J Clin Iimmunol. 2010;30(5):693–702. doi:10.1007/s10875-010-9425-2
82. Xu H, Yang J, Gao W, et al. Innate immune sensing of bacterial modifications of Rho GTPases by the Pyrin inflammasome. Nature. 2014;513(7517):237–241. doi:10.1038/nature13449
83. Ting JP, Lovering RC, Alnemri ES, et al. The NLR gene family: a standard nomenclature. Immunity. 2008;28(3):285–287. doi:10.1016/j.immuni.2008.02.005
84. Huang M, Zhang X. Structural and biochemical mechanisms of NLRP1 inhibition by DPP9. Nature. 2021;592(7856):773–777. doi:10.1038/s41586-021-03320-w
85. Sandstrom A, Mitchell PS, Goers L, Mu EW. Functional degradation: a mechanism of NLRP1 inflammasome activation by diverse pathogen enzymes. Science (New York, N.Y.). 2019;364(6435). doi:10.1126/science.aau1330
86. Mitchell PS, Sandstrom A, Vance RE. The NLRP1 inflammasome: new mechanistic insights and unresolved mysteries. Current Opinion in Immunology. 2019;60:37–45. doi:10.1016/j.coi.2019.04.015
87. Kahlenberg JM, Dubyak GR. Mechanisms of caspase-1 activation by P2X7 receptor-mediated K+ release. Am J Physiol Cell Physiol. 2004;286(5):C1100–1108. doi:10.1152/ajpcell.00494.2003
88. Zhao Y, Shao F. The NAIP-NLRC4 inflammasome in innate immune detection of bacterial flagellin and type III secretion apparatus. Immunol Rev. 2015;265(1):85–102. doi:10.1111/imr.12293
89. Hornung V, Ablasser A, Charrel-Dennis M, et al. AIM2 recognizes cytosolic dsDNA and forms a caspase-1-activating inflammasome with ASC. Nature. 2009;458(7237):514–518. doi:10.1038/nature07725
90. Aubert DF, Xu H, Yang J, et al. A Burkholderia Type VI Effector Deamidates Rho GTPases to Activate the Pyrin Inflammasome and Trigger Inflammation. Cell Host Microbe. 2016;19(5):664–674. doi:10.1016/j.chom.2016.04.004
91. Smith C, Wang X, Yin H. Caspases come together over LPS. Trend Immunol. 2015;36(2):59–61. doi:10.1016/j.it.2014.12.007
92. Chu LH, Indramohan M, Ratsimandresy RA, et al. The oxidized phospholipid oxPAPC protects from septic shock by targeting the non-canonical inflammasome in macrophages. Nat Commun. 2018;9(1):996. doi:10.1038/s41467-018-03409-3
93. Aglietti RA, Dueber EC. Recent Insights into the Molecular Mechanisms Underlying Pyroptosis and Gasdermin Family Functions. Trend Immunol. 2017;38(4):261–271. doi:10.1016/j.it.2017.01.003
94. Wang K, Sun Q, Zhong X, et al. Structural Mechanism for GSDMD Targeting by Autoprocessed Caspases in Pyroptosis. Cell. 2020;180(5):941–955.e920. doi:10.1016/j.cell.2020.02.002
95. Kelley N, Jeltema D, Duan Y, He Y. The NLRP3 Inflammasome: an Overview of Mechanisms of Activation and Regulation. Int J Mol Sci. 2019;20(13):1.
96. Zheng X, Wan J, Tan G. The mechanisms of NLRP3 inflammasome/pyroptosis activation and their role in diabetic retinopathy. Front Immunol. 2023;14:1151185. doi:10.3389/fimmu.2023.1151185
97. Yang D, He Y, Muñoz-Planillo R, Liu Q, Núñez G. Caspase-11 Requires the Pannexin-1 Channel and the Purinergic P2X7 Pore to Mediate Pyroptosis and Endotoxic Shock. Immunity. 2015;43(5):923–932. doi:10.1016/j.immuni.2015.10.009
98. He Y, Zeng MY, Yang D, Motro B, Núñez G. NEK7 is an essential mediator of NLRP3 activation downstream of potassium efflux. Nature. 2016;530(7590):354–357. doi:10.1038/nature16959
99. Crespo yanguas S, Willebrords J, Johnstone SR, et al. Pannexin1 as mediator of inflammation and cell death. Biochim Biophys Acta Mol Cell Res. 2017;1864(1):51–61. doi:10.1016/j.bbamcr.2016.10.006
100. Kayagaki N, Warming S, Lamkanfi M, et al. Non-canonical inflammasome activation targets caspase-11. Nature. 2011;479(7371):117–121. doi:10.1038/nature10558
101. Downs KP, Nguyen H, Dorfleutner A, Stehlik C. An overview of the non-canonical inflammasome. Mol Aspect Med. 2020;76:100924. doi:10.1016/j.mam.2020.100924
102. Zhang CC, Li CG, Wang YF, et al. Chemotherapeutic paclitaxel and cisplatin differentially induce pyroptosis in A549 lung cancer cells via caspase-3/GSDME activation. Apoptosis: an International Journal on Programmed Cell Death. 2019;24(3–4):312–325. doi:10.1007/s10495-019-01515-1
103. Orning P, Weng D, Starheim K, Ratner D. Pathogen blockade of TAK1 triggers caspase-8-dependent cleavage of gasdermin D and cell death. Science (New York, N.Y.). 2018;362(6418):1064–1069. doi:10.1126/science.aau2818
104. Sarhan J, Liu BC, Muendlein HI, et al. Caspase-8 induces cleavage of gasdermin D to elicit pyroptosis during Yersinia infection. Proc Natl Acad Sci USA. 2018;115(46):E10888–e10897. doi:10.1073/pnas.1809548115
105. Zheng Z, Deng W, Bai Y, et al. The Lysosomal Rag-Ragulator Complex Licenses RIPK1 and Caspase-8-mediated Pyroptosis by Yersinia. Science. 2021;372(6549). doi:10.1126/science.abg0269.
106. Hou J, Zhao R, Xia W, et al. PD-L1-mediated gasdermin C expression switches apoptosis to pyroptosis in cancer cells and facilitates tumour necrosis. Nature Cell Biology. 2020;22(10):1264–1275. doi:10.1038/s41556-020-0575-z
107. Shi J, Zhao Y, Wang Y, et al. Inflammatory caspases are innate immune receptors for intracellular LPS. Nature. 2014;514(7521):187–192. doi:10.1038/nature13683
108. Zheng M, Kanneganti TD. Newly Identified Function of Caspase-6 in ZBP1-mediated Innate Immune Responses, NLRP3 Inflammasome Activation, PANoptosis, and Host Defense. J Cell Iimmunol. 2020;2(6):341–347. doi:10.33696/immunology.2.064
109. Liu Y, Fang Y. Gasdermin E-mediated target cell pyroptosis by CAR T cells triggers cytokine release syndrome. Science Immunol. 2020;5(43):7969.
110. Zhang Z, Zhang Y, Xia S, et al. Gasdermin E suppresses tumour growth by activating anti-tumour immunity. Nature. 2020;579(7799):415–420. doi:10.1038/s41586-020-2071-9
111. Zhong X, Zeng H, Zhou Z, Su Y, Cheng H, Hou Y. Structural mechanisms for regulation of GSDMB pore-forming activity. Nature. 2023;616(7957):598–605. doi:10.1038/s41586-023-05872-5
112. Zhou Z, He H. Granzyme A from cytotoxic lymphocytes cleaves GSDMB to trigger pyroptosis in target cells. Science. 2020;368(6494):7548.
113. Vafaei S, Taheri H, Hajimomeni Y, Fakhre Yaseri A, Abolhasani Zadeh F. The role of NLRP3 inflammasome in colorectal cancer: potential therapeutic target. Clinical & Translational Oncology: Official Publication of the Federation of Spanish Oncology Societies and of the National Cancer Institute of Mexico. 2022;24(10):1881–1889. doi:10.1007/s12094-022-02861-4
114. Zhang X, Li C, Chen D, et al. H. pylori CagA activates the NLRP3 inflammasome to promote gastric cancer cell migration and invasion. Inflamm Res. 2022;71(1):141–155. doi:10.1007/s00011-021-01522-6
115. Wu X, Dong L, Lin X, Li J. Relevance of the NLRP3 Inflammasome in the Pathogenesis of Chronic Liver Disease. Front Immunol. 2017;8:1728. doi:10.3389/fimmu.2017.01728
116. Nagarajan K, Soundarapandian K, Thorne RF, Li D, Li D. Activation of Pyroptotic Cell Death Pathways in Cancer: an Alternative Therapeutic Approach. Transl Oncol. 2019;12(7):925–931. doi:10.1016/j.tranon.2019.04.010
117. Zhou W, Liu H, Yuan Z, et al. Targeting the mevalonate pathway suppresses ARID1A-inactivated cancers by promoting pyroptosis. Cancer Cell. 2023;41(4):740–756.e10. doi:10.1016/j.ccell.2023.03.002
118. van den Heerik A, Horeweg N. Adjuvant therapy for endometrial cancer in the era of molecular classification: radiotherapy, chemoradiation and novel targets for therapy. International Journal of Gynecological Cancer: Official Journal of the International Gynecological Cancer Society. 2021;31(4):594–604. doi:10.1136/ijgc-2020-001822
119. Chai R, Li Y, Shui L, Ni L, Zhang A. The role of pyroptosis in inflammatory diseases. Front Cell Develop Biol. 2023;11:1173235. doi:10.3389/fcell.2023.1173235
120. Yang Y, Liu PY, Bao W, Chen SJ, Wu FS, Zhu PY. Hydrogen inhibits endometrial cancer growth via a ROS/NLRP3/caspase-1/GSDMD-mediated pyroptotic pathway. BMC Cancer. 2020;20(1):28. doi:10.1186/s12885-019-6491-6
121. Murakami M, Osuka S. Effectiveness of NLRP3 Inhibitor as a Non-Hormonal Treatment for ovarian endometriosis. Reproductive Biology and Endocrinology: RB&E. 2022;20(1):58. doi:10.1186/s12958-022-00924-3
122. Rébé C, Ghiringhelli F. Interleukin-1β and Cancer. Cancers. 2020;12(7):1791.
123. Lopez-Castejon G, Brough D. Understanding the mechanism of IL-1β secretion. Cytokine Growth Factor Rev. 2011;22(4):189–195. doi:10.1016/j.cytogfr.2011.10.001
124. Martín-Sánchez F, Diamond C, Zeitler M, et al. Inflammasome-dependent IL-1β release depends upon membrane permeabilisation. Cell Death & Differentiation. 2016;23(7):1219–1231. doi:10.1038/cdd.2015.176
125. Abais JM, Xia M, Zhang Y, Boini KM, Li PL. Redox regulation of NLRP3 inflammasomes: ROS as trigger or effector? Antioxid Redox Signal. 2015;22(13):1111–1129. doi:10.1089/ars.2014.5994
126. Tian K, Yang Y, Zhou K, et al. The role of ROS-induced pyroptosis in CVD. Front Cardiovasc Med. 2023;10:1116509. doi:10.3389/fcvm.2023.1116509
127. Kim SM, Hwang KA, Choi KC. Potential roles of reactive oxygen species derived from chemical substances involved in cancer development in the female reproductive system. BMB Rep. 2018;51(11):557–562. doi:10.5483/BMBRep.2018.51.11.056
128. Jiang Y, Wang M, Huang K, et al. Oxidized low-density lipoprotein induces secretion of interleukin-1β by macrophages via reactive oxygen species-dependent NLRP3 inflammasome activation. Biochem Biophys Res Commun. 2012;425(2):121–126. doi:10.1016/j.bbrc.2012.07.011
129. Villalpando-Rodriguez GE, Gibson SB. Reactive Oxygen Species (ROS) Regulates Different Types of Cell Death by Acting as a Rheostat. Oxidative Medicine and Cellular Longevity. 2021;2021:9912436. doi:10.1155/2021/9912436
130. Milkovic L, Cipak Gasparovic A, Cindric M, Mouthuy PA. Short Overview of ROS as Cell Function Regulators and Their Implications in Therapy Concepts. Cells. 2019;8(8):793.
131. Checa J, Aran JM. Reactive Oxygen Species: drivers of Physiological and Pathological Processes. J Inflamm Res. 2020;13:1057–1073. doi:10.2147/JIR.S275595
132. Redza-Dutordoir M, Averill-Bates DA. Activation of apoptosis signalling pathways by reactive oxygen species. BBA. 2016;1863(12):2977–2992. doi:10.1016/j.bbamcr.2016.09.012
133. Ren JD, Wu XB, Jiang R, Hao DP, Liu Y. Molecular hydrogen inhibits lipopolysaccharide-triggered NLRP3 inflammasome activation in macrophages by targeting the mitochondrial reactive oxygen species. BBA. 2016;1863(1):50–55. doi:10.1016/j.bbamcr.2015.10.012
134. Auten RL, Davis JM. Oxygen toxicity and reactive oxygen species: the devil is in the details. Pediatr Res. 2009;66(2):121–127. doi:10.1203/PDR.0b013e3181a9eafb
135. Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem Biol Interact. 2006;160(1):1–40. doi:10.1016/j.cbi.2005.12.009
136. Zhu X, Xu Y, Wang J, Xue Z, Qiu T, Chen J. Loss of NLRP3 reduces oxidative stress and polarizes intratumor macrophages to attenuate immune attack on endometrial cancer. Front Immunol. 2023;14:1165602. doi:10.3389/fimmu.2023.1165602
137. Katturajan R, Nithiyanandam S, Parthasarathy M, et al. Immunomodulatory Role of Thioredoxin Interacting Protein in Cancer’s Impediments: current Understanding and Therapeutic Implications. Vaccines. 2022;10(11):1902. doi:10.3390/vaccines10111902
138. Masutani H. Thioredoxin-Interacting Protein in Cancer and Diabetes. Antioxid Redox Signal. 2022;36(13–15):1001–1022. doi:10.1089/ars.2021.0038
139. Yang C, Xia W, Liu X, Lin J, Wu A. Role of TXNIP/NLRP3 in sepsis-induced myocardial dysfunction. IntJ Mol Med. 2019;44(2):417–426. doi:10.3892/ijmm.2019.4232
140. Tsubaki H, Tooyama I. Thioredoxin-Interacting Protein (TXNIP) with Focus on Brain and Neurodegenerative Diseases. Int J Mol Sci. 2020;21(24):9357. doi:10.3390/ijms21249357
141. Deng J, Pan T, Liu Z, et al. The role of TXNIP in cancer: a fine balance between redox, metabolic, and immunological tumor control. Br J Cancer. 2023;129(12):1877–1892. doi:10.1038/s41416-023-02442-4
142. Park JW, Lee SH, Woo GH, Kwon HJ, Kim DY. Downregulation of TXNIP leads to high proliferative activity and estrogen-dependent cell growth in breast cancer. Biochemical and Biophysical Research Communications. 2018;498(3):566–572. (). doi:10.1016/j.bbrc.2018.03.020
143. Hu J, Feng L, Ren M, et al. Colorectal Cancer Cell Differentiation Is Dependent on the Repression of Aerobic Glycolysis by NDRG2-TXNIP Axis. Digestive Diseases and Sciences. 2022;67(8):3763–3772. doi:10.1007/s10620-021-07188-8
144. Lim JY, Yoon S, Hong S, et al. Thioredoxin and thioredoxin-interacting protein as prognostic markers for gastric cancer recurrence. World Journal of Gastroenterology. 2012;18(39):5581. (). doi:10.3748/wjg.v18.i39.5581
145. Kim Y, Kim YS, Kim M, Kim JM, Lee HH, Kim TH. Thioredoxin-interacting Protein (TXNIP) Mediates Thioredoxin-dependent Antioxidant Mechanism in Endometrial Cancer Cells Treated With 1α,25-dihydroxyvitamin D(3). Anticancer Res. 2019;39(9):4795–4803. doi:10.21873/anticanres.13664
146. Chen Y, Ning J, Cao W, et al. Research Progress of TXNIP as a Tumor Suppressor Gene Participating in the Metabolic Reprogramming and Oxidative Stress of Cancer Cells in Various Cancers. Front Oncol. 2020;10:568574. doi:10.3389/fonc.2020.568574
147. Gunes A, Bagirsakci E, Iscan E, et al. Thioredoxin interacting protein promotes invasion in hepatocellular carcinoma. Oncotarget. 2018;9(96):36849–36866. doi:10.18632/oncotarget.26402
148. Iqbal MA, Chattopadhyay S, Siddiqui FA, et al. Silibinin induces metabolic crisis in triple-negative breast cancer cells by modulating EGFR-MYC-TXNIP axis: potential therapeutic implications. FEBS J. 2021;288(2):471–485. doi:10.1111/febs.15353
149. Li Y, Miao LY, Xiao YL, et al. Hypoxia induced high expression of thioredoxin interacting protein (TXNIP) in non-small cell lung cancer and its prognostic effect. APJCP. 2015;16(7):2953–2958. doi:10.7314/APJCP.2015.16.7.2953
150. Li J, Yue Z, Xiong W, Sun P, You K, Wang J. TXNIP overexpression suppresses proliferation and induces apoptosis in SMMC7221 cells through ROS generation and MAPK pathway activation. Oncology Reports. 2017;37(6):3369–3376. (). doi:10.3892/or.2017.5577
151. Lin L, Zhang MX, Zhang L, Zhang D, Li C, Autophagy LYL. Pyroptosis, and Ferroptosis: new Regulatory Mechanisms for Atherosclerosis. Front Cell Develop Biol. 2021;9:809955. doi:10.3389/fcell.2021.809955
152. Zheng Z, Li G. Mechanisms and Therapeutic Regulation of Pyroptosis in Inflammatory Diseases and Cancer. Int J Mol Sci. 2020;21(4):1456. doi:10.3390/ijms21041456
153. Wang J, Wang Y. Novel Risk Classification Based on Pyroptosis-Related Genes Defines Immune Microenvironment and Pharmaceutical Landscape for Hepatocellular Carcinoma. Cancers. 2022;14(2):447.
154. Huang X, Li Y, Li J, Yang X, Xiao J, Xu F. The Expression of Pyroptosis-Related Gene May Influence the Occurrence, Development, and Prognosis of Uterine Corpus Endometrial Carcinoma. Front Oncol. 2022;12:885114. doi:10.3389/fonc.2022.885114
155. Yu C, Qi H, Zhang Y, Zhao W, Wu G. Elevated Expression of Gamma-Glutamyl Hydrolase Is Associated With Poor Prognosis and Altered Immune Signature in Uterine Corpus Endometrial Carcinoma. Front Genetics. 2021;12:764194. doi:10.3389/fgene.2021.764194
156. Zheng X, Lv X, Zhu L, et al. The Circadian Gene NPAS2 Act as a Putative Tumor Stimulative Factor for Uterine Corpus Endometrial Carcinoma. Cancer Manage Res. 2021;13:9329–9343. doi:10.2147/CMAR.S343097
157. Liu J, Geng R, Yang S, et al. Development and Clinical Validation of Novel 8-Gene Prognostic Signature Associated With the Proportion of Regulatory T Cells by Weighted Gene Co-Expression Network Analysis in Uterine Corpus Endometrial Carcinoma. Front Immunol. 2021;12:788431. doi:10.3389/fimmu.2021.788431
158. Li Y, Wang Y, Guo H, Wu Q, Hu Y. IRF2 contributes to myocardial infarction via regulation of GSDMD induced pyroptosis. Mol Med Rep. 2022;25(2):1.
159. Xia X, Wang X, Cheng Z, et al. The role of pyroptosis in cancer: pro-cancer or pro-”host”? Cell Death Dis. 2019;10(9):650. doi:10.1038/s41419-019-1883-8
160. Wei X, Xie F, Zhou X, et al. Role of pyroptosis in inflammation and cancer. Cellular & Molecular Immunology. 2022;19(9):971–992. doi:10.1038/s41423-022-00905-x
161. Qiu S, Hu Y, Dong S. Pan-cancer analysis reveals the expression, genetic alteration and prognosis of pyroptosis key gene GSDMD. Int Immunopharmacol. 2021;101(Pt A):108270. doi:10.1016/j.intimp.2021.108270
162. Li L, Li Y, Bai Y. Role of GSDMB in Pyroptosis and Cancer. Cancer Management and Research. 2020;12:3033–3043. doi:10.2147/CMAR.S246948
163. Liang H, Van Remmen H, Frohlich V, Lechleiter J, Richardson A, Ran Q. Gpx4 protects mitochondrial ATP generation against oxidative damage. Biochem Biophys Res Commun. 2007;356(4):893–898. doi:10.1016/j.bbrc.2007.03.045
164. Dabkowski ER, Williamson CL, Hollander JM. Mitochondria-specific transgenic overexpression of phospholipid hydroperoxide glutathione peroxidase (GPx4) attenuates ischemia/reperfusion-associated cardiac dysfunction. Free Radic Biol Med. 2008;45(6):855–865. doi:10.1016/j.freeradbiomed.2008.06.021
165. Yang Y, Yang L, Jiang S, et al. HMGB1 mediates lipopolysaccharide-induced inflammation via interacting with GPX4 in colon cancer cells. Cancer Cell International. 2020;20:205. doi:10.1186/s12935-020-01289-6
166. Lee N, Carlisle AE, Peppers A, Park SJ, Doshi MB. xCT-Driven Expression of GPX4 Determines Sensitivity of Breast Cancer Cells to Ferroptosis Inducers. Antioxidant. 2021;10(2):1.
167. Guerriero E, Capone F, Accardo M, et al. GPX4 and GPX7 over-expression in human hepatocellular carcinoma tissues. EJH. 2015;59(4):2540. doi:10.4081/ejh.2015.2540
168. Shao R, Wang X, Xu T, Xia Y, Cui D. The balance between AIM2-associated inflammation and autophagy: the role of CHMP2A in brain injury after cardiac arrest. Journal of Neuroinflammation. 2021;18(1):257. doi:10.1186/s12974-021-02307-8
169. Rosadini CV, Kagan JC. Early innate immune responses to bacterial LPS. Current Opinion in Immunology. 2017;44:14–19. doi:10.1016/j.coi.2016.10.005
170. Zhang G, Ghosh S. Toll-like receptor-mediated NF-kappaB activation: a phylogenetically conserved paradigm in innate immunity. J Clin Invest. 2001;107(1):13–19. doi:10.1172/JCI11837
171. Li X, Feng M, Zhao Y, et al. A Novel TLR4-Binding Domain of Peroxiredoxin From Entamoeba histolytica Triggers NLRP3 Inflammasome Activation in Macrophages. Front Immunol. 2021;12:758451. doi:10.3389/fimmu.2021.758451
172. Zhang Y, Yang W, Li W, Zhao Y. NLRP3 Inflammasome: checkpoint Connecting Innate and Adaptive Immunity in Autoimmune Diseases. Front Immunol. 2021;12:732933. doi:10.3389/fimmu.2021.732933
173. Kayagaki N, Wong MT, Stowe IB, et al. Noncanonical inflammasome activation by intracellular LPS independent of TLR4. Science. 2013;341(6151):1246–1249. doi:10.1126/science.1240248
174. Demon D, Vande Walle L, Lamkanfi M. Sensing the enemy within: how macrophages detect intracellular Gram-negative bacteria. Trends Biochem Sci. 2014;39(12):574–576. doi:10.1016/j.tibs.2014.10.006
175. Hagar JA, Powell DA, Aachoui Y, Ernst RK, Miao EA. Cytoplasmic LPS activates caspase-11: implications in TLR4-independent endotoxic shock. Science. 2013;341(6151):1250–1253. doi:10.1126/science.1240988
176. Rathinam VAK, Zhao Y, Shao FA-O. Innate immunity to intracellular LPS. Nature Immunology. 2019;20(5):527–533. (). doi:10.1038/s41590-019-0368-3
177. Li W, Deng M, Loughran PA, et al. LPS Induces Active HMGB1 Release From Hepatocytes Into Exosomes Through the Coordinated Activities of TLR4 and Caspase-11/GSDMD Signaling. Frontiers in Immunology. 2020;11:. doi:10.3389/fimmu.2020.00229
178. Savio LEB, de Andrade Mello P, da Silva CG, Coutinho-Silva R. The P2X7 Receptor in Inflammatory Diseases: angel or Demon? Front Pharmacol. 2018;9:52.
179. Karmakar M, Katsnelson MA, Dubyak GA-O, Pearlman E. Neutrophil P2X7 receptors mediate NLRP3 inflammasome-dependent IL-1β secretion in response to ATP. Nature Communications. 2016;7(1):. doi:10.1038/ncomms10555.
180. Mariathasan S, Weiss Ds Fau - Newton K, Newton K, et al. Cryopyrin activates the inflammasome in response to toxins and ATP. Nature. 2006;440(7081):228–232. doi:10.1038/nature04515
181. Yang Y, Wang H, Kouadir M, Song H, Shi F Recent advances in the mechanisms of NLRP3 inflammasome activation and its inhibitors. Cell Death Dis. 2019;10:128.
182. Latz E, Xiao Ts Fau - Stutz TS, Stutz A. Activation and regulation of the inflammasomes. Nature Reviews Immunology. 2013;13(6):397–411. (). doi:10.1038/nri3452
183. Zhang L, Ai C, Bai M, Niu J, Zhang Z. NLRP3 Inflammasome/Pyroptosis: a Key Driving Force in Diabetic Cardiomyopathy. International Journal of Molecular Sciences. 2022;23(18):10632. doi:10.3390/ijms231810632.
184. Li X, Qi X, Fau - Zhou L, et al. P2X(7) receptor expression is decreased in epithelial cancer cells of ectodermal, uro-genital sinus, and distal paramesonephric duct origin. Purinergic Signalling. 2009;5:351.
185. Li X, Qi X, Zhou L, et al. P2X(7) receptor expression is decreased in epithelial cancer cells of ectodermal, uro-genital sinus, and distal paramesonephric duct origin. Purinergic Signalling. 2009;5(3):351–368. doi:10.1007/s11302-009-9161-3
186. Bulun SE, Yilmaz BD, Sison C, et al. Endometriosis. Endocrine Rev. 2019;40(4):1048–1079. doi:10.1210/er.2018-00242
187. Smolarz B, Szyłło K. Endometriosis: epidemiology, Classification, Pathogenesis, Treatment and Genetics (Review of Literature). Int J Mol Sci. 2021;22(19):10554. doi:10.3390/ijms221910554
188. Xu Y, Liu H, Xiong W, et al. A novel mechanism regulating pyroptosis-induced fibrosis in endometriosis via lnc-MALAT1/miR-141-3p/NLRP3 pathway†. Biology of reproduction. Biology of Reproduction. 2023;109(2):156–171. doi:10.1093/biolre/ioad057
189. Fonseca BM, Pinto B, Costa L, Felgueira E, Rebelo I. Increased expression of NLRP3 inflammasome components in granulosa cells and follicular fluid interleukin(IL)-1beta and IL-18 levels in fresh IVF/ICSI cycles in women with endometriosis. Journal of Assisted Reproduction and Genetics. 2023;40(1):191–199. doi:10.1007/s10815-022-02662-2
190. Bullon P, Navarro JM. Inflammasome as a Key Pathogenic Mechanism in Endometriosis. Current Drug Targets. 2017;18(9):997–1002. doi:10.2174/1389450117666160709013850
191. Zhou F, Zhao F, Huang Q, Lin X, Zhang S, Dai Y. NLRP3 activated macrophages promote endometrial stromal cells migration in endometriosis. J Reprod Immunol. 2022;152:103649. doi:10.1016/j.jri.2022.103649
192. Zhang M, Shi Z, Peng X, et al. NLRP3 inflammasome-mediated Pyroptosis induce Notch signal activation in endometriosis angiogenesis. Mol Cell Endocrinol. 2023;574:111952. doi:10.1016/j.mce.2023.111952
193. Hang Y, Tan L, Chen Q, Liu Q, Jin Y. E3 ubiquitin ligase TRIM24 deficiency promotes NLRP3/caspase-1/IL-1β-mediated pyroptosis in endometriosis. Cell Biol Int. 2021;45(7):1561–1570. doi:10.1002/cbin.11592
194. Zhao J, Ma W, Chen W, et al. AEG-1 aggravates inflammation via promoting NALP3 inflammasome formation in murine endometriosis lesions. Anim Cells Syst. 2019;23(6):407–413. doi:10.1080/19768354.2019.1691052
195. Nakamura Y, Kambe N. Linkage of bacterial colonization of skin and the urticaria-like rash of NLRP3-mediated autoinflammatory syndromes through mast cell-derived TNF-α. J Dermatological Sci. 2013;71(2):83–88. doi:10.1016/j.jdermsci.2013.04.009
196. Mangan MSJ, Olhava EJ, Roush WR, Seidel HM, Glick GD, Latz E. Targeting the NLRP3 inflammasome in inflammatory diseases. Nat Rev Drug Discov. 2018;17(8):588–606. doi:10.1038/nrd.2018.97
197. Guo X, Xu X, Li T, et al. NLRP3 Inflammasome Activation of Mast Cells by Estrogen via the Nuclear-Initiated Signaling Pathway Contributes to the Development of Endometriosis. Front Immunol. 2021;12:749979. doi:10.3389/fimmu.2021.749979
198. Wilhite AM, Baca Y, Xiu J, et al. Molecular profiles of endometrial cancer tumors among Black patients. Gynecol Oncol. 2022;166(1):108–116. doi:10.1016/j.ygyno.2022.04.014
199. Cuccu I, D’Oria O, Sgamba L, et al. Role of Genomic and Molecular Biology in the Modulation of the Treatment of Endometrial Cancer: narrative Review and Perspectives. Healthcare. 2023;11(4). doi:10.3390/healthcare11040571.
200. Henry CE, Phan K, Orsman EJ, Kenwright D, Thunders MC, Filoche SK. Molecular Profiling of Endometrial Cancer: an Exploratory Study in Aotearoa, New Zealand. Cancers. 2021;13(22):5641. doi:10.3390/cancers13225641
201. Kim J, Kim SI, Kim NR, et al. Prognostic significance of L1CAM expression in addition to ProMisE in endometrial cancer. Gynecol Oncol. 2023;174:231–238. doi:10.1016/j.ygyno.2023.05.062
202. Kommoss FK, Karnezis AN, Kommoss F, et al. L1CAM further stratifies endometrial carcinoma patients with no specific molecular risk profile. Br J Cancer. 2018;119(4):480–486. doi:10.1038/s41416-018-0187-6
203. Giannini A, D’Oria O, Corrado G, et al. The role of L1CAM as predictor of poor prognosis in stage I endometrial cancer: a systematic review and meta-analysis. Arch Gynecol Obstet. 2024;309(3):789–799. doi:10.1007/s00404-023-07149-8
204. Haruma T, Nagasaka T, Nakamura K, et al. Clinical impact of endometrial cancer stratified by genetic mutational profiles, POLE mutation, and microsatellite instability. PLoS One. 2018;13(4):e0195655. doi:10.1371/journal.pone.0195655
205. Bosse T, Nout RA, Stelloo E, et al. L1 cell adhesion molecule is a strong predictor for distant recurrence and overall survival in early stage endometrial cancer: pooled PORTEC trial results. Eur J Cancer. 2014;50(15):2602–2610. doi:10.1016/j.ejca.2014.07.014
206. Nakao A, Toyoda Y, Sharma P, Evans M, Guthrie N. Effectiveness of hydrogen rich water on antioxidant status of subjects with potential metabolic syndrome-an open label pilot study. J Clin Biochem Nutr. 2010;46(2):140–149. doi:10.3164/jcbn.09-100
207. Nagatani K, Nawashiro H, Takeuchi S, et al. Safety of intravenous administration of hydrogen-enriched fluid in patients with acute cerebral ischemia: initial clinical studies. Med Gas Res. 2013;3(1):13. doi:10.1186/2045-9912-3-13
208. Liu Y, Yang L, Tao K, et al. Protective effects of hydrogen enriched saline on liver ischemia reperfusion injury by reducing oxidative stress and HMGB1 release. BMC Gastroenterol. 2014;14(1):12. doi:10.1186/1471-230X-14-12
209. Zhao P, Jin Z, Chen Q, et al. Local generation of hydrogen for enhanced photothermal therapy. Nat Commun. 2018;9(1):4241. doi:10.1038/s41467-018-06630-2
210. Xie F, Song Y, Yi Y, et al. Therapeutic Potential of Molecular Hydrogen in Metabolic Diseases from Bench to Bedside. Pharmaceuticals. 2023;16(4):1.
211. Meng J, Liu L, Wang D, Yan Z, Chen G. Hydrogen gas represses the progression of lung cancer via down-regulating CD47. Biosci Rep. 2020;40(4). doi:10.1042/BSR20192761
212. Asgharzadeh F, Tarnava A, Mostafapour A, Khazaei M, LeBaron TW. Hydrogen-rich water exerts anti-tumor effects comparable to 5-fluorouracil in a colorectal cancer xenograft model. World Journal of Gastrointestinal Oncology. 2022;14(1):242–252. doi:10.4251/wjgo.v14.i1.242
213. Chen JB, Lu YY, Xu KC. A narrative review of hydrogen oncology: from real world survey to real world evidence. Med Gas Res. 2020;10(3):130–133. doi:10.4103/2045-9912.296044
214. Zhao L, Zhou C, Zhang J, et al. Hydrogen protects mice from radiation induced thymic lymphoma in BALB/c mice. Int J Bio Sci. 2011;7(3):297–300. doi:10.7150/ijbs.7.297
215. Yang Y, Zhu Y, Xi X. Anti-inflammatory and antitumor action of hydrogen via reactive oxygen species. Oncol Lett. 2018;16(3):2771–2776. doi:10.3892/ol.2018.9023
216. Kitamura A, Kobayashi S, Matsushita T, Fujinawa H, Murase K. Experimental verification of protective effect of hydrogen-rich water against cisplatin-induced nephrotoxicity in rats using dynamic contrast-enhanced CT. Br J Radiol. 2010;83(990):509–514. doi:10.1259/bjr/25604811
217. Malik A, Kanneganti TD. Inflammasome activation and assembly at a glance. Journal of Cell Science. 2017;130(23):3955–3963. doi:10.1242/jcs.207365
218. Liston A, Masters SL. Homeostasis-altering molecular processes as mechanisms of inflammasome activation. Nat Rev Immunol. 2017;17(3):208–214. doi:10.1038/nri.2016.151
219. Han SJ, Jung SY, Wu SP, et al. Estrogen Receptor β Modulates Apoptosis Complexes and the Inflammasome to Drive the Pathogenesis of Endometriosis. Cell. 2015;163(4):960–974. doi:10.1016/j.cell.2015.10.034
220. Dawson A, Fernandez ML, Anglesio M, Yong PJ, Carey MS. Endometriosis and endometriosis-associated cancers: new insights into the molecular mechanisms of ovarian cancer development. Ecancermedicalscience. 2018;12:803. doi:10.3332/ecancer.2018.803
221. Brilhante AV, Augusto KL, Portela MC, et al. Endometriosis and Ovarian Cancer: an Integrative Review (Endometriosis and Ovarian Cancer). APJCP. 2017;18(1):11–16. doi:10.22034/APJCP.2017.18.1.11
222. Liu H, Wan J, Chu J. Long non-coding RNAs and endometrial cancer. Biomed Pharmacothe. 2019;119:109396. doi:10.1016/j.biopha.2019.109396
223. Vallone C, Rigon G, Gulia C, et al. Non-Coding RNAs and Endometrial Cancer. Genes. 2018;9(4). doi:10.3390/genes9040187.
224. Aljubran F, Nothnick WB. Long non-coding RNAs in endometrial physiology and pathophysiology. Mol Cell Endocrinol. 2021;525:111190. doi:10.1016/j.mce.2021.111190
225. He WP, Chen YY, Wu LX, Guo YY, You ZS, Yang GF. A novel necroptosis-related lncRNA signature for predicting prognosis and anti-cancer treatment response in endometrial cancer. Front Immunol. 2022;13:1018544. doi:10.3389/fimmu.2022.1018544
226. Kim LK, Park SA. LncRNA SNHG4 Modulates EMT Signal and Antitumor Effects in Endometrial Cancer through Transcription Factor SP-1. Biomedicines. 2023;11(4). doi:10.3390/biomedicines11041018
227. Tian J, Cheng H, Wang N, Wang C. SLERT, as a novel biomarker, orchestrates endometrial cancer metastasis via regulation of BDNF/TRKB signaling. World J Surgl Oncol. 2023;21(1):27. doi:10.1186/s12957-022-02821-w
228. Shan S, Wang X, Qian L, Wang C, Zhao S. ENST00000534735 inhibits proliferation and migration, promotes apoptosis and pyroptosis of endometrial cancer via OSBPL3 through APMK/SIRT1/NF-κB pathway. Heliyon. 2024;10(4):e25281. doi:10.1016/j.heliyon.2024.e25281
229. Mao X, Lei H, Yi T, et al. Lipid reprogramming induced by the TFEB-ERRα axis enhanced membrane fluidity to promote EC progression. Journal of Experimental & Clinical Cancer Research: CR. 2022;41(1):28. doi:10.1186/s13046-021-02211-2
230. Ma L, Zong X. Metabolic Symbiosis in Chemoresistance: refocusing the Role of Aerobic Glycolysis. Front Oncol. 2020;10:5. doi:10.3389/fonc.2020.00005
231. Zeng B, Chen Y, Chen H, et al. Exosomal miR-211-5p regulates glucose metabolism, pyroptosis, and immune microenvironment of melanoma through GNA15. Pharmacol Res. 2023;188:106660. doi:10.1016/j.phrs.2023.106660
232. Catanzaro D, Gaude E, Orso G, et al. Inhibition of glucose-6-phosphate dehydrogenase sensitizes cisplatin-resistant cells to death. Oncotarget. 2015;6(30):30102–30114. doi:10.18632/oncotarget.4945
233. Feng J, Dai W, Mao Y, et al. Simvastatin re-sensitizes hepatocellular carcinoma cells to sorafenib by inhibiting HIF-1α/PPAR-γ/PKM2-mediated glycolysis. Journal of Experimental & Clinical Cancer Research: CR. 2020;39(1):24. doi:10.1186/s13046-020-1528-x
234. He Q, Hao P, He G, et al. IGF2BP1-regulated expression of ERRα is involved in metabolic reprogramming of chemotherapy resistant osteosarcoma cells. J Transl Med. 2022;20(1):348. doi:10.1186/s12967-022-03549-7
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.