")
Back to Journals » Journal of Inflammation Research » Volume 17
Roles of Microenvironment on Mesenchymal Stem Cells Therapy for Osteoarthritis
Authors Zhang H, Jin C, Hua J, Chen Z , Gao W, Xu W, Zhou L , Shan L
Received 25 April 2024
Accepted for publication 21 September 2024
Published 3 October 2024 Volume 2024:17 Pages 7069—7079
DOI https://doi.org/10.2147/JIR.S475617
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Haiyan Zhang,1,* Chaoying Jin,2,* Jiaqing Hua,3,* Zuxiang Chen,4 Wenxin Gao,4 Wenting Xu,4 Li Zhou,4 Letian Shan1
1The Second Affiliated Hospital, Zhejiang Chinese Medical University, Hangzhou, Zhejiang, People’s Republic of China; 2School of Medicine, Zhejiang University, Hangzhou, Zhejiang, People’s Republic of China; 3College of Pharmaceutical Science, Zhejiang Chinese Medical University, Hangzhou, Zhejiang, People’s Republic of China; 4The First Affiliated Hospital, Zhejiang Chinese Medical University, Hangzhou, Zhejiang, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Letian Shan; Li Zhou, Email [email protected]; [email protected]
Abstract: Osteoarthritis (OA) induced microenvironmental alterations are a common and unavoidable phenomenon that greatly exacerbate the pathologic process of OA. Imbalances in the synthesis and degradation of cartilage extracellular matrix (ECM) have been reported to be associated with an adverse microenvironment. Stem cell therapy is a promising treatment for OA, and mesenchymal stem cells (MSCs) are the main cell sources for this therapy. With multispectral differentiation and immunomodulation, MSCs can effectively regulate the microenvironment of articular cartilage, ameliorate inflammation, promote regeneration of damaged cartilage, and ultimately alleviate OA symptoms. However, the efficacy of MSCs in the treatment of OA is greatly influenced by articular cavity microenvironments. This article reviews the five microenvironments of OA articular cavity, including inflammatory microenvironment, senescence microenvironment, hypoxic microenvironment, high glucose microenvironment and high lipid environment, focus on the positive and negative effects of OA microenvironments on the fate of MSCs. In this regard, we emphasize the mechanisms of the current use of MSCs in OA treatment, as well as its limitations and challenges.
Keywords: microenvironment, mesenchymal stem cells, osteoarthritis
Introduction
Articular cartilage contains a variety of cell types, consisting of chondrocytes, synoviocytes, endothelial cells, mesenchymal stem cells (MSCs), and immune cells.1,2 These cellular components form a normal articular cavity microenvironment and maintain articular cartilage homeostasis.2 In pathological states, unfavorable transformation undergoes in the articular microenvironment. The oxidative stress balance in chondrocytes and subchondral bone is destroyed, and the synthesis and degradation of extracellular matrix (ECM) are disrupted, leading to the occurrence of osteoarthritis (OA).3,4
OA is the leading cause of disability in the older people without radical treatments.5,6 In recent years, MSCs therapy is proved to be a promising therapy for OA treatment. MSCs can effectively regulate the microenvironment of articular cavity, ameliorate inflammation, promote cartilage regeneration and repair, and ultimately alleviate the symptoms of OA.7,8
MSCs are a class of pluripotent stem cells, which originate in the mesoderm and have the capacity for self-renewal and multilineage differentiation, including chondrogenic, osteogenic, and adipogenic potential.9 Besides, MSCs also have immunomodulatory, homing, and anti-inflammatory properties,10 which support their versatility as bone tissue therapeutic agents.11 The efficacy, safety and reliability of MSCs therapy in OA treatment has been demonstrated in numerous clinical trials and preclinical studies.12–14 However, the efficacy of MSCs is affected by the microenvironment of the articular cavity, and hostile articular microenvironment can impair the bioactivity of MSCs.15 Therefore, altering the adverse articular cavity microenvironment may be an effective strategy for OA treatment.16
The microenvironment in which stem cells live is also called as the stem cell niche. It maintains critical stem cell properties. This microenvironment contains a variety of elements from systemic and localized sources, including cellular and ECM components.17 Within the ecological niche, stem cells can interact with other cells as well as the ECM, releasing soluble signals that regulate stem cells function, thereby maintaining tissue homeostasis and promoting damage repair.18 In brief, the stem cell niche provides a special microenvironment for stem cells, enabling them to remain in an undifferentiated state. Factors released by cells in the stem cell niche direct the stem cells fate by activating relevant intracellular signaling pathways.19 However, the microenvironment of OA lesions deviates greatly from the stem cell niche under physiological conditions, exerting considerable impacts on the local therapeutic effects of stem cells. Therefore, research on such impact is also an important basis for us to improve stem cell treatment methods.
This paper reviews the properties and roles of MSCs and the relationship between MSCs and the microenvironment of OA, focusing on the positive and negative effects of the five microenvironments of the OA articular cavity (inflammatory microenvironment, senescence microenvironment, hypoxic microenvironment, high glucose microenvironment and high lipid microenvironment) on MSCs fate. The mechanisms of MSCs as a treatment for OA and their interactions and challenges are highlighted (Figure 1).
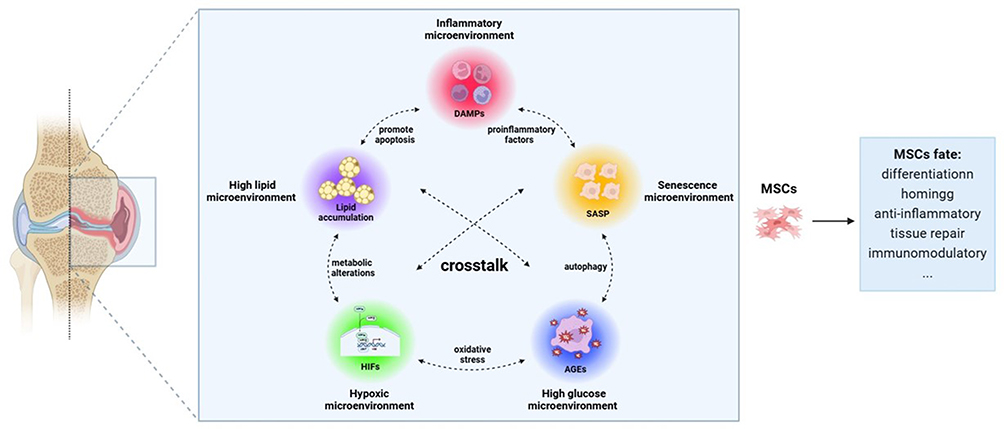 |
Figure 1 Regulation of MSCs fate by OA microenvironment. Notes: Inflammatory microenvironment, senescence microenvironment, hypoxic microenvironment, high glucose microenvironment, and high lipid microenvironment in OA articular cavity alter the fate of MSCs and affect the therapeutic effect of MSCs. |
Inflammatory Microenvironment
Accumulating evidence suggests that chronic low-level inflammation contributes to the accelerated the progression of OA.20–22 Such process involves activation of the innate immune response.23,24 In OA microenvironment, pro-inflammatory cytokines induce degradation of the ECM. The accumulated degraded products or cartilage fragments are considered to be damage-associated molecular patterns (DAMPs). DAMPs activate pattern-recognition receptors (PRRs), including Toll-like receptors (TLRs), NOD-like receptors (NLRs), and receptor for advanced glycosylation end-products (RAGEs), the innate immune system is then activated.25,26 PRRs stimulate fibroblast-like synoviocytes (FLS), macrophages, and chondrocytes to produce and release various pro-inflammatory factors into synovial fluid. This results in loss of chondrocyte phenotype and ECM degradation. Ultimately, an inflammatory microenvironment conducive to cartilage disease is formed.27,28
Immune Cells in the Inflammatory Microenvironment
Under physiological conditions, macrophages maintain microenvironmental homeostasis by phagocytosis of pathogens and debris in senescent tissues, whereas in OA articular cavity, abnormally activated macrophages may exacerbate joint destruction.29 Macrophages have two main polarization states: M1 macrophages and M2 macrophages.30 Macrophage polarization to phenotype-1 macrophages enhances the pathological process of OA by driving inflammation, whereas phenotype-2 macrophages are anti-inflammatory and facilitate rapid tissue repair.31,32 It has been found that mediators released by damaged chondrocytes provide a pro-inflammatory microenvironment to promote M1 polarization.33 M1 macrophages can produce interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), matrix metalloproteinases (MMPs), and other inflammatory cytokines, which release matrix metalloproteinases to promote the degradation of cartilage matrix.34 Neutrophils arrive at the injury site immediately after OA injury and form a local neutrophilic infiltration in the joint.35,36 The progression of OA is also associated with the expansion of pathogenic T lymphocytes and MSCs have been shown to successfully prevent T cells proliferation.37,38
MSCs in the Inflammatory Microenvironment
MSCs inhibit the inflammatory microenvironment by decreasing pro-inflammatory factors and inhibiting B cells infiltration.39 Through influencing the interaction of B lymphocytes and T lymphocytes, MSCs influence the adaptive immune system.30 Specifically, MSCs can directly bind to the PD-9 receptor on B cells, thereby preventing B cells from activating into plasma cells.40 Moreover, MSCs can also upregulate PD-1 receptors on Tregs to enhance Tregs-mediated immunosuppression.41 The anti-inflammatory cytokine IL-10 secreted by MSCs can induce the transformation of macrophages from M1 to M2, mediating anti-inflammatory effects.42 MSCs exert immunosuppressive effects by converting pro-inflammatory M1 macrophages into anti-inflammatory M2 macrophages, thereby attenuating the macrophage-induced inflammatory microenvironment.43 MSCs or MSCs conditioned medium significantly reduced the number of infiltrating neutrophils and alleviated the progression of OA.44
Inflammatory Microenvironment and MSCs Fate
The inflammatory microenvironment in which MSCs reside affects their self-renewal, multilineage differentiation and proliferation capabilities.45 The Wnt/β-catenin pathway regulates the differentiation of MSCs.46 In inflammatory microenvironment, mitochondrial autophagy is inhibited by activating Wnt/β-catenin pathway. Thus, damaged mitochondria accumulate continuously in MSCs and impair their differentiation.47 Inflammatory factors in the inflammatory microenvironment activate the Wnt/β-catenin pathway, leading to decreased differentiation ability of MSCs.48 In inflammatory microenvironment, the cartilage protective effect of human umbilical cord mesenchymal stem cells (hucMSCs) was weakened, which impaired the repair effect of hucMSCs15 (Figure 2).
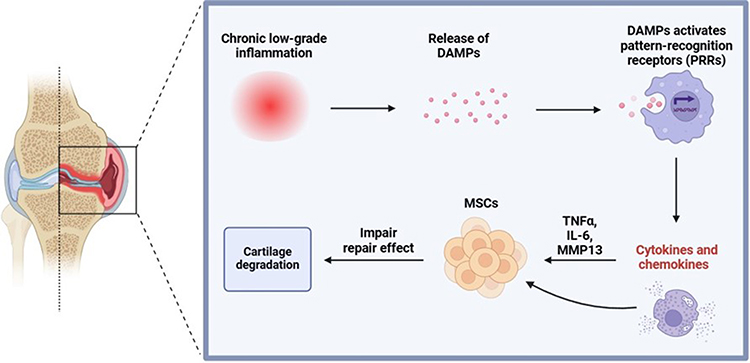 |
Figure 2 The inflammatory microenvironment in OA. Notes: Chronic low-grade inflammation releases DAMPs, activates PRRs to release pro-inflammatory factors to impair the cartilage repair effect of MSCs. |
Although most of the evidence suggests the adverse effects of the inflammatory microenvironment on MSCs, the inflammatory microenvironment also enhances the anti-inflammatory effects of MSCs. MSCs exert their biological effects by releasing exosomes.49 For example, IL-1β-induced MSCs exosomes significantly enhanced their anti-inflammatory activity in osteoarthritic cells SW982 compared to MSC-Exo.50 In a double-damage rabbit OA model, intraarticular injection of human adipose-derived stem cells (hADSCs) reduced joint effusion, which helped to attenuate the inflammatory microenvironment in the joint cavity and promote cartilage repair.51 In vitro inflammatory exposure enhances the immunomodulatory capacity of MSCs, and the resulting secretome can protect chondrocytes from catabolism.52 Hypoxia and inflammatory microenvironment can protect MSCs from damage in harsh microenvironments, thereby increasing the survival, homing, and paracrine capacities of MSCs in vitro and in vivo.53 Inflammation and lesion sites trigger enhanced MSC homing for tissue remodeling and repair.54,55
Overall, the different fates of MSC in the inflammatory microenvironment may be related to their status.
Senescence Microenvironment
The process of organismal senescence is accompanied by cellular senescence, which caused by a sustained DNA damage response, telomere dysfunction, stem cells exhaustion, and impaired intercellular communication.56 Senescent cells accumulate in osteoarthritic joints and release factors called senescence-associated secretory phenotype (SASP), which include the pro-inflammatory cytokine IL-6 and IL-8, monocyte chemotactic protein 1 (MCP1), and ECM proteases.57,58 The transcriptional activation of a SASP program affects the microenvironment in which senescent cells reside and reinforces cellular senescence.59 SASP can accelerate cell senescence and deteriorate the neighboring cells function.56
Senescence Microenvironment and MSCs Fate
MSCs are critical cells for maintaining tissue homeostasis and organization during tissue and organ injury.60 The exhaustion of stem cells is the marker of aging.61 Senescent MSCs are able to function in the surrounding environment and regulate MSCs niche.62 The functional capacity and their number of MSCs declines with age.63,64 During aging, MSCs experience functional degeneration and gradual loss of stemness, with subsequent changes in the fate of MSCs and aberrant differentiation.65 Accumulation of senescent MSCs in the articular cavity may aggravate the process of OA.66 Senescence OA-MSCs cause senescence, inflammation and fibrosis in cartilage.67 Studies have found that MSCs isolated from OA patients are influenced by aging-driven OA inflammation and nutritional microenvironment, resulting in diminished cartilage anabolism and enhanced cartilage catabolism.68 Recent studies have shown that injection of senescent MSCs in the joint cavity of young mice drives an OA-like phenotype.69 In the senescence microenvironment, MSCs lose stemness and inhibit chondrogenic differentiation, and the paracrine role of MSCs is also affected.69 Compared with MSCs from young donors, senescent donor-expanded MSCs exhibit increased expression of senescence markers, elevated levels of inflammation, decreased immunoregulatory function, and reduced proliferation and differentiation.70,71 Aging MSCs contribute to the establishment and maintenance of an inflammatory environment in stem cell niche. In senescent MSCs, SASP effect factor IL-1α and IL-8 induce early senescence of MSCs in a NF-κb dependent manner through paracrine.72 There is a correlation between MSCs aging and the age-related disease OA, which is accelerated by enhanced m6A levels and down-regulated expression of the demethylation enzyme ALKBH5 during MSCs aging.73
Hypoxic Microenvironment
Due to the lack of blood supply, articular cartilage is located in a hypoxic environment throughout life.74 Hypoxia-inducible factor (HIF) is a core factor in inducing hypoxia gene and regulating cellular oxygen environment, mediating chondrocyte adaptation to the hypoxic microenvironment.75 HIFs are heterodimers of heterogeneous proteins polymerized from two distinct subunits, the oxygen-sensitively regulated α subunit (HIF-1α, HIF-2α & HIF-3α) and the structurally expressed HIF-1β subunit.76 The most important members of the HIFs protein family are HIF-1α and HIF-2α. In hypoxic environment, HIF-1α undergoes activation and enters the nucleus, where it binds to the subunit to form an active HIF-1α transcription factors, which are involved in autophagy and apoptosis, promotion of chondrocyte phenotype and maintenance of chondrocyte viability.77 Unlike HIF-1α, HIF-2α is mainly involved in promoting the expression of chondrocytes catabolic factors, up-regulates the expression of the apoptotic gene Fas, increasing chondrocyte apoptosis, and exacerbating inflammatory responses, leading to more severe articular cartilage destruction.78
Hypoxic Microenvironment and MSCs Fate
In the past, it was thought that hypoxia is beneficial for MSC, numerous studies have shown that cultured under hypoxic conditions would exhibit enhanced MSCs proliferative potential and stemness.79–81 Since the oxygen concentration of tissue is less than 5%, cell culture of MSCs under hypoxic environment can mimic in vivo microenvironment and help to maintain the physiological processes of MSCs from differentiation, proliferation, and metabolic homeostasis.82 Hypoxia can induce upregulation of pluripotent markers of MSCs, enabling MSCs to maintain their function.83 Hypoxic microenvironment maintains the undifferentiated phenotype of MSCs.84 In addition, HIF acts as the regulatory molecule of the hypoxic microenvironment to control the differentiation and fate of MSCs.85 Isolation and amplification of bone marrow mesenchymal stem cells (BMSCs) under hypoxic environments can up-regulate the expression of chondrogenesis genes and enhance the chondrogenic differentiation of BMSCs.86 Different oxygen concentrations had different effects on adipose tissue-derived MSCs (ADSCs).87 In vitro, hypoxic preconditioning of MSCs can up-regulate the expression of MSCs chemokine receptors CXCR4, CXCR7 and CX3CR1, which contributes to the migration and tissue repair of MSCs.88
Despite the prevailing view that hypoxia is beneficial to stem cells, it can also adversely affect MSCs. Chronic hypoxia predisposes BMSCs from patients with cyanotic congenital heart disease (CCHD) to premature senescence, possibly due to dysbiosis of the intestinal flora, which induces the accumulation of D-galactose.89 It was shown that hypoxic microenvironment in osteonecrosis zone led to hypoxia-induced apoptosis in transplanted BMSCs. Under hypoxic condition, the expression of the pro-apoptotic gene Bax was increased, whereas down-regulation expression of apoptosis inhibitor Bcl-2, leading to apoptosis in more than 70% of BMSCs.90 Notably, severe hypoxia inhibits the adhesion and mobility of BMSCs, which limits their use.91 The hypoxic microenvironment also leads to increased cellular oxidative stress, leading to senescence of MSCs and affecting MSCs differentiation.92,93
High Glucose Microenvironment
Obesity and the related disease have been shown to be risk factors for developing OA.94 Diabetes mellitus (DM) is an obesity-related disease, which is characterized by hyperglycemia, which chronically affects the physiological function of the cells involved and leads to their damage.95 High blood glucose levels in patients are involved in a variety of complex pathologic events, including a rise in ROS, cellular stress, and inflammatory cytokines.96,97 Hyperglycemia also induces the production and accumulation of advanced glycosylation end products (AGEs), which adversely affects and irreversibly alters the microenvironment in which the cells live.
High Glucose Microenvironment and MSCs Fate
MSC-based cell therapy treatments are promising for treating diabetes, but the microenvironmental stress in patients with hyperglycemia can significantly inhibit the survival time of MSCs in human bodies, which ultimately affects their therapeutic efficacy. In vitro, the high glycemic microenvironment also affected the culture efficiency of MSCs.
In diabetic patients, the number of circulating MSCs is reduced, causing an insufficient number of MSCs migrate to the injury site, and the regeneration and repair of target tissue is impaired.98 The diabetic microenvironment impaired the therapeutic effect of MSCs infusion on bone reduction, which is attributed to the fact that the hyperglycemic condition inhibited MSC-T-cell interactions through the AMPK pathway and reduced the anti-inflammatory capacity of MSCs.99 The high glucose microenvironment down-regulated the AKT-Sirt1-TWIST pathway and inhibited TWIST exacerbated high glucose-induced apoptosis in MSCs but promoted osteogenic differentiation of MSCs.100 A recent study showed that hAD-MSCs cultured in high glucose had reduced immunosuppressive capacity, which was associated with significant reductions in the levels of IDO, IL-10, and complement factor levels, as well as a significant increase in immune proteasome activity.95 Exposure of AGEs to the diabetic environment leads to stem cell dysfunction. The increase of AGEs inhibits the proliferation of MSCs, induces apoptosis of MSCs and prevents the differentiation of MSCs to adipose, cartilage, and bone.101 AGE-BSA induces chemokine/cytokine production through activation of ROS-p38-mediated pathways. These chemokines/cytokines have inhibiting effects on the growth and migration of MSCs.102 In most cases, the articular cavity microenvironments are crosstalk with each other. High glucose induces BMSCs to produce ROS through activating NADPH oxidase. ROS activates autophagy by upregulating ATGs expression and induces BMSCs senescence.103 High glucose and hypoxia upregulate HIF-1α expression, which promotes inflammation and oxidative stress.104 In stable state, MSCs depend on glycolysis, have low ROS content, and high ROS levels can damage proteins and DNA at the telomere level, inducing replicative aging, also known as stress-induced premature aging.105 These results suggest that the articular cavity microenvironments are much more complex than previously thought.
High Lipid Microenvironment
Excessive lipid accumulation is a hallmark of obesity-related pathologies.106 Abnormally high lipid levels promote the release of inflammatory mediators and induce macrophage polarization toward the inflammatory M1 phenotype, causing an inflammatory response in the body.107 Previous studies have suggested that obesity promotes excessive lipid deposition in non-adipose tissues, which leads to lipotoxicity and tissue dysfunction.108 Recent evidence suggests that articular cartilage accumulates lipids during the aging process, and lipid accumulation in cartilage was found to be positively correlated with the severity of knee OA (kOA).109
High Lipid Microenvironment and MSCs Fate
Abnormal lipid levels in vivo interfere with MSCs function, affect MSCs homing, and inhibit tissue regeneration.110 Multispectral differentiation capacity of MSCs is key to their use as clinical regenerative therapy for kOA. Studies have shown an inverse correlation between adipogenesis and osteogenesis in MSCs. The presence of fat may increase adipocyte proliferation, differentiation, and fat accumulation while inhibiting osteoblast differentiation and bone formation.111 The lipid-lowering agent lovastatin inhibits adipogenesis and stimulates osteogenic differentiation by inhibiting peroxisome proliferator-activated receptor γ2 (PPARγ2) and increasing Runx2 expression in BMSCs.112
Although lipid accumulation affects multiple functions of stem cells, exosomes secreted by MSCs also play a role in inhibiting lipid accumulation. HucMSCs attenuate injury associated with lipid deposition, collagen deposition, or inflammation in vivo and in vitro.113 HucMSCs-derived exosomes ameliorate inflammation, abnormal lipid accumulation, and mitochondrial oxidative stress in non-alcoholic steatohepatitis (NASH) mice.114 Notably, MSCs survival, proliferation, and terminal differentiation are associated with ROS regulation, and ROS affects MSCs adipogenesis or osteogenic differentiation by modulating the MSCs differentiation signaling cascade response.115 Obesity is an important cause of the formation of a high glucose and high lipid microenvironment, and the accumulation of excess lipids in diabetic bone tissue causes changes in the bone marrow microenvironment, which reprograms bone metabolism.116 Moreover, intermittent hypoxia is an independent risk factor for metabolic dysfunction-associated fatty liver disease (MAFLD).
Conclusion and Discussion
This paper reviews five kinds of articular cavity microenvironments that affect the fate of MSCs. MSCs play a therapeutic role in damaged articular cartilage, including ameliorate inflammation, chondrogenic differentiation, proliferation, migration and tissue repair. As shown in Table 1, the microenvironment of the articular cavity largely determines the fate of MSCs. A healthy articular cavity microenvironment can regulate the proliferation and differentiation of MSCs, enabling MSCs to function normally in immunomodulation and tissue repair. In a diseased articular cavity microenvironment, the function of MSCs is compromised and instead aggravates the process of OA.117
 |
Table 1 Fate of MSCs in Different Microenvironments |
In fact, the articular cavity microenvironments are crosstalk with each other, and the mechanism of determining the fate of MSCs by the microenvironment is very complex, which is need more research to change the articular cavity microenvironment to make it more suitable for the survival of MSCs. For the time being, organoids may be useful in helping to study the effects of the microenvironment on MSCs. Organoids are self-assembled 3D structures in vitro, mainly generated from primitive tissues or MSCs, that can be used to mimic the complex microenvironments corresponding to organs.118 The articular cavity microenvironments of different OA patients vary to a large extent, and the development of 3D organoids can help personalize treatment for different patients and achieve precision medicine.118 As a clinical disease, OA’s joint microenvironment is far more complex than we imagined. According to individual differences in patients’ microenvironment, treatment methods also need to be personalized. By detecting the joint fluid of OA patients and analyzing what kind of microenvironment the joint cavity is in, it is helpful to realize the symptomatic treatment of OA for different joint cavity microenvironments in the future.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Natural Science Foundation of China (grant numbers 82274373, 82104890, and 82074464), Zhejiang Traditional Chinese Medical Science Foundation (grant number 2022ZB137), and General Research Project of Education Department of Zhejiang Province (grant number Y202248740).
Disclosure
The authors declare that they have no competing interests in this work.
References
1. You B, Zhou C, Yang Y. MSC-EVs alleviate osteoarthritis by regulating microenvironmental cells in the articular cavity and maintaining cartilage matrix homeostasis. Ageing Res Rev. 2023;85:101864. doi:10.1016/j.arr.2023.101864
2. Wu Y, Li J, Zeng Y, et al. Exosomes rewire the cartilage microenvironment in osteoarthritis: from intercellular communication to therapeutic strategies. Int J Oral Sci. 2022;14(1):40. doi:10.1038/s41368-022-00187-z
3. Ahmad N, Ansari MY, Haqqi TM. Role of iNOS in osteoarthritis: pathological and therapeutic aspects. J Cell Physiol. 2020;235(10):6366–6376. doi:10.1002/jcp.29607
4. Hou W, Ye C, Chen M, et al. Excavating bioactivities of nanozyme to remodel microenvironment for protecting chondrocytes and delaying osteoarthritis. Bioact Mater. 2021;6(8):2439–2451. doi:10.1016/j.bioactmat.2021.01.016
5. Mianehsaz E, Mirzaei HR, Mahjoubin-Tehran M, et al. Mesenchymal stem cell-derived exosomes: a new therapeutic approach to osteoarthritis? Stem Cell Res Ther. 2019;10(1):340. doi:10.1186/s13287-019-1445-0
6. Zheng L, Zhang Z, Sheng P, Mobasheri A. The role of metabolism in chondrocyte dysfunction and the progression of osteoarthritis. Ageing Res Rev. 2021;66:101249. doi:10.1016/j.arr.2020.101249
7. Xu X, Xu L, Xia J, Wen C, Liang Y, Zhang Y. Harnessing knee joint resident mesenchymal stem cells in cartilage tissue engineering. Acta Biomater. 2023;168:372–387. doi:10.1016/j.actbio.2023.07.024
8. Carneiro DC, Araújo LT, Santos GC, et al. Clinical Trials with Mesenchymal Stem Cell Therapies for Osteoarthritis: challenges in the Regeneration of Articular Cartilage. Int J Mol Sci. 2023;24(12). doi:10.3390/ijms24129939
9. Yang Y, Peng Y, Li Y, Shi T, Luan Y, Yin C. Role of stem cell derivatives in inflammatory diseases. Front Immunol. 2023;14:1153901. doi:10.3389/fimmu.2023.1153901
10. Alfaro D, Rodríguez-Sosa MR, Zapata AG, et al. Eph/ephrin Signaling and Biology of Mesenchymal Stromal/Stem Cells. Journal of clinical medicine. 2020;9(2):310. doi:10.3390/jcm9020310
11. Liu X, Fang T, Shi T, Wang Y, Liu G. Hydrogels provide microenvironments to mesenchymal stem cells for craniofacial bone regeneration: review. J Biomater Appl. 2023;38(1):3–24. doi:10.1177/08853282231183103
12. Lamo-Espinosa JM, Mora G, Blanco JF, et al. Intra-articular injection of two different doses of autologous bone marrow mesenchymal stem cells versus hyaluronic acid in the treatment of knee osteoarthritis: long-term follow up of a multicenter randomized controlled clinical trial (phase I/II). J Transl Med. 2018;16(1):213. doi:10.1186/s12967-018-1591-7
13. Copp G, Robb KP, Viswanathan S. Culture-expanded mesenchymal stromal cell therapy: does it work in knee osteoarthritis? A pathway to clinical success. Cell Mol Immunol. 2023;20(6):626–650. doi:10.1038/s41423-023-01020-1
14. Liu TP, Ha P, Xiao CY, et al. Updates on mesenchymal stem cell therapies for articular cartilage regeneration in large animal models. Front Cell Develop Biol. 2022;10:982199. doi:10.3389/fcell.2022.982199
15. Fan M, Tong P, Yan L, et al. Detrimental alteration of mesenchymal stem cells by an articular inflammatory microenvironment results in deterioration of osteoarthritis. BMC Med. 2023;21(1):215. doi:10.1186/s12916-023-02923-6
16. Lepetsos P, Papavassiliou KA, Papavassiliou AG. Redox and NF-κB signaling in osteoarthritis. Free Radic Biol Med. 2019;132:90–100. doi:10.1016/j.freeradbiomed.2018.09.025
17. Farahzadi R, Valipour B, Montazersaheb S, Fathi E. Targeting the stem cell niche micro-environment as therapeutic strategies in aging. Front Cell Develop Biol. 2023;11:1162136. doi:10.3389/fcell.2023.1162136
18. Pennings S, Liu KJ, Qian H. The Stem Cell Niche: interactions between Stem Cells and Their Environment. Stem Cells Int. 2018;2018:4879379. doi:10.1155/2018/4879379
19. Chacón-Martínez CA, Koester J, Wickström SA. Signaling in the stem cell niche: regulating cell fate, function and plasticity. Development. 2018;145(15). doi:10.1242/dev.165399
20. De Roover A, Escribano-Núñez A, Monteagudo S, Lories R. Fundamentals of osteoarthritis: inflammatory mediators in osteoarthritis. Osteoarthritis Cartilage. 2023;31(10):1303–1311. doi:10.1016/j.joca.2023.06.005
21. Terkawi MA, Ebata T, Yokota S, et al. Low-Grade Inflammation in the Pathogenesis of Osteoarthritis: cellular and Molecular Mechanisms and Strategies for Future Therapeutic Intervention. Biomedicines. 2022;10(5):1109. doi:10.3390/biomedicines10051109
22. Partain BD, Bracho-Sanchez E, Farhadi SA, et al. Intra-articular delivery of an indoleamine 2,3-dioxygenase galectin-3 fusion protein for osteoarthritis treatment in male Lewis rats. Arthritis Res Therapy. 2023;25(1):173. doi:10.1186/s13075-023-03153-0
23. Pérez-García S, Calamia V, Hermida-Gómez T, et al. Proteomic Analysis of Synovial Fibroblasts and Articular Chondrocytes Co-Cultures Reveals Valuable VIP-Modulated Inflammatory and Degradative Proteins in Osteoarthritis. Int J Mol Sci. 2021;22(12):6441. doi:10.3390/ijms22126441
24. Bartels YL, van Lent P, van der Kraan PM, Blom AB, Bonger KM, van den Bosch MHJ. Inhibition of TLR4 signalling to dampen joint inflammation in osteoarthritis. Rheumatology. 2023;63(3):608–618. doi:10.1093/rheumatology/kead493
25. Chung HY, Kim DH, Lee EK, et al. Redefining Chronic Inflammation in Aging and Age-Related Diseases: proposal of the Senoinflammation Concept. Aging Dis. 2019;10(2):367–382. doi:10.14336/ad.2018.0324
26. Lambert C, Zappia J, Sanchez C, Florin A, Dubuc JE, Henrotin Y. The Damage-Associated Molecular Patterns (DAMPs) as Potential Targets to Treat Osteoarthritis: perspectives From a Review of the Literature. Front Med. 2020;7:607186. doi:10.3389/fmed.2020.607186
27. Rahmati M, Mobasheri A, Mozafari M. Inflammatory mediators in osteoarthritis: a critical review of the state-of-The-art, current prospects, and future challenges. Bone. 2016;85:81–90. doi:10.1016/j.bone.2016.01.019
28. Semenistaja S, Skuja S, Kadisa A, Groma V. Healthy and Osteoarthritis-Affected Joints Facing the Cellular Crosstalk. Int J Mol Sci. 2023;24(4):4120. doi:10.3390/ijms24044120
29. Zhang H, Lin C, Zeng C, et al. Synovial macrophage M1 polarisation exacerbates experimental osteoarthritis partially through R-spondin-2. Ann Rheumatic Dis. 2018;77(10):1524–1534. doi:10.1136/annrheumdis-2018-213450
30. Rizzo MG, Best TM, Huard J, et al. Therapeutic Perspectives for Inflammation and Senescence in Osteoarthritis Using Mesenchymal Stem Cells, Mesenchymal Stem Cell-Derived Extracellular Vesicles and Senolytic Agents. Cells. 2023;12(10):1421. doi:10.3390/cells12101421
31. Lv Z, Xu X, Sun Z, et al. TRPV1 alleviates osteoarthritis by inhibiting M1 macrophage polarization via Ca(2+)/CaMKII/Nrf2 signaling pathway. Cell Death Dis. 2021;12(6):504. doi:10.1038/s41419-021-03792-8
32. Chen Y, Yang L, Li X. Advances in Mesenchymal stem cells regulating macrophage polarization and treatment of sepsis-induced liver injury. Front Immunol. 2023;14:1238972. doi:10.3389/fimmu.2023.1238972
33. Kulkarni P, Srivastava V, Tootsi K, et al. Synovial Fluid in Knee Osteoarthritis Extends Proinflammatory Niche for Macrophage Polarization. Cells. 2022;11(24):4115. doi:10.3390/cells11244115
34. Shapouri-Moghaddam A, Mohammadian S, Vazini H, et al. Macrophage plasticity, polarization, and function in health and disease. J Cell Physiol. 2018;233(9):6425–6440. doi:10.1002/jcp.26429
35. Huang L, Dong G, Peng J, et al. The role of exosomes and their enhancement strategies in the treatment of osteoarthritis. Human Cell. 2023;36(6):1887–1900. doi:10.1007/s13577-023-00970-y
36. Ribitsch I, Bileck A, Egerbacher M, et al. Fetal Immunomodulatory Environment Following Cartilage Injury-The Key to CARTILAGE Regeneration? Int J Mol Sci. 2021;22(23):12969. doi:10.3390/ijms222312969
37. Krampera M, Glennie S, Dyson J, et al. Bone marrow mesenchymal stem cells inhibit the response of naive and memory antigen-specific T cells to their cognate peptide. Blood. 2003;101(9):3722–3729. doi:10.1182/blood-2002-07-2104
38. Ribeiro A, Laranjeira P, Mendes S, et al. Mesenchymal stem cells from umbilical cord matrix, adipose tissue and bone marrow exhibit different capability to suppress peripheral blood B, natural killer and T cells. Stem Cell Res Ther. 2013;4(5):125. doi:10.1186/scrt336
39. Rockel JS, Rabani R, Viswanathan S. Anti-fibrotic mechanisms of exogenously-expanded mesenchymal stromal cells for fibrotic diseases. Semin Cell Dev Biol. 2020;101:87–103. doi:10.1016/j.semcdb.2019.10.014
40. Klein-Wieringa IR, de Lange-Brokaar BJ, Yusuf E, et al. Inflammatory Cells in Patients with Endstage Knee Osteoarthritis: a Comparison between the Synovium and the Infrapatellar Fat Pad. J Rheumatol. 2016;43(4):771–778. doi:10.3899/jrheum.151068
41. Yan Z, Zhuansun Y, Chen R, Li J, Ran P. Immunomodulation of mesenchymal stromal cells on regulatory T cells and its possible mechanism. Exp Cell Res. 2014;324(1):65–74. doi:10.1016/j.yexcr.2014.03.013
42. Yadav P, Singh SK, Rajput S, et al. Therapeutic potential of stem cells in regeneration of liver in chronic liver diseases: current perspectives and future challenges. Pharmacol Ther. 2023;253:108563. doi:10.1016/j.pharmthera.2023.108563
43. Tian R, Su S, Yu Y, et al. Revolutionizing osteoarthritis treatment: how mesenchymal stem cells hold the key. Biomed Pharmacothe. 2024;173:116458. doi:10.1016/j.biopha.2024.116458
44. Wang G, Cao K, Liu K, et al. Kynurenic acid, an IDO metabolite, controls TSG-6-mediated immunosuppression of human mesenchymal stem cells. Cell Death Differ. 2018;25(7):1209–1223. doi:10.1038/s41418-017-0006-2
45. Xiao J, Gong X, Fu Z, et al. The influence of inflammation on the characteristics of adipose-derived mesenchymal stem cells (ADMSCs) and tissue repair capability in a hepatic injury mouse model. Stem Cell Res Ther. 2023;14(1):334. doi:10.1186/s13287-023-03532-z
46. Schizas NP, Zafeiris C, Neri AA, Anastasopoulos PP, Papaioannou NA, Dontas IA. Inhibition versus activation of canonical Wnt-signaling, to promote chondrogenic differentiation of Mesenchymal Stem Cells. A review. Orthoped Rev. 2021;13(2):27098. doi:10.52965/001c.27098
47. Zhai Q, Chen X, Fei D, et al. Nanorepairers Rescue Inflammation-Induced Mitochondrial Dysfunction in Mesenchymal Stem Cells. Adv Sci. 2022;9(4):e2103839. doi:10.1002/advs.202103839
48. Alongi DJ, Yamaza T, Song Y, et al. Stem/progenitor cells from inflamed human dental pulp retain tissue regeneration potential. Regenerat Med. 2010;5(4):617–631. doi:10.2217/rme.10.30
49. Li Q, Li B, Ye T, et al. Requirements for human mesenchymal stem cell‐derived small extracellular vesicles. Interdiscip Med. 2023;1(1):e20220015. doi:10.1002/INMD.20220015
50. Kim M, Shin DI, Choi BH, Min BH. Exosomes from IL-1β-Primed Mesenchymal Stem Cells Inhibited IL-1β- and TNF-α-Mediated Inflammatory Responses in Osteoarthritic SW982 Cells. Tissue Eng and Regener Med. 2021;18(4):525–536. doi:10.1007/s13770-020-00324-x
51. Wang J, Su S, Dong C, et al. Human Adipose-derived Stem Cells Upregulate IGF-1 and Alleviate Osteoarthritis in a Two-stage Rabbit Osteoarthritis Model. Curr Stem Cell Res Ther. 2024;2024:1. doi:10.2174/011574888x274359231122064109
52. Cassano JM, Schnabel LV, Goodale MB, Fortier LA. Inflammatory licensed equine MSCs are chondroprotective and exhibit enhanced immunomodulation in an inflammatory environment. Stem Cell Res Ther. 2018;9(1):82. doi:10.1186/s13287-018-0840-2
53. Hu C, Wu Z, Li L. Pre-treatments enhance the therapeutic effects of mesenchymal stem cells in liver diseases. J Cell Mol Med. 2020;24(1):40–49. doi:10.1111/jcmm.14788
54. Kidd S, Spaeth E, Klopp A, Andreeff M, Hall B, Marini FC. The (in) auspicious role of mesenchymal stromal cells in cancer: be it friend or foe. Cytotherapy. 2008;10(7):657–667. doi:10.1080/14653240802486517
55. Mishra VK, Shih HH, Parveen F, et al. Identifying the Therapeutic Significance of Mesenchymal Stem Cells. Cells. 2020;9(5):1145. doi:10.3390/cells9051145
56. Di Micco R, Krizhanovsky V, Baker D, d’Adda Di Fagagna F. Cellular senescence in ageing: from mechanisms to therapeutic opportunities. Nat Rev Mol Cell Biol. 2021;22(2):75–95. doi:10.1038/s41580-020-00314-w
57. van Deursen JM. The role of senescent cells in ageing. Nature. 2014;509(7501):439–446. doi:10.1038/nature13193
58. Acosta JC, Banito A, Wuestefeld T, et al. A complex secretory program orchestrated by the inflammasome controls paracrine senescence. Nat Cell Biol. 2013;15(8):978–990. doi:10.1038/ncb2784
59. Georget M, Defois A, Guiho R, et al. Development of a DNA damage-induced senescence model in osteoarthritic chondrocytes. Aging. 2023;15(17):8576–8593. doi:10.18632/aging.204881
60. Williams AR, Hare JM. Mesenchymal stem cells: biology, pathophysiology, translational findings, and therapeutic implications for cardiac disease. Circ Res. 2011;109(8):923–940. doi:10.1161/circresaha.111.243147
61. López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013;153(6):1194–1217. doi:10.1016/j.cell.2013.05.039
62. Coppé JP, Patil CK, Rodier F, et al. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008;6(12):2853–2868. doi:10.1371/journal.pbio.0060301
63. Warren LA, Rossi DJ. Stem cells and aging in the hematopoietic system. Mechani Ageing Development. 2009;130(1–2):46–53. doi:10.1016/j.mad.2008.03.010
64. Yu KR, Kang KS. Aging-related genes in mesenchymal stem cells: a mini-review. Gerontology. 2013;59(6):557–563. doi:10.1159/000353857
65. Weng Z, Wang Y, Ouchi T, et al. Mesenchymal Stem/Stromal Cell Senescence: hallmarks, Mechanisms, and Combating Strategies. Stem Cells Transl Med. 2022;11(4):356–371. doi:10.1093/stcltm/szac004
66. Fu L, Hu Y, Song M, et al. Up-regulation of FOXD1 by YAP alleviates senescence and osteoarthritis. PLoS Biol. 2019;17(4):e3000201. doi:10.1371/journal.pbio.3000201
67. Liu W, Brodsky AS, Feng M, et al. Senescent Tissue-Resident Mesenchymal Stromal Cells Are an Internal Source of Inflammation in Human Osteoarthritic Cartilage. Front Cell Develop Biol. 2021;9:725071. doi:10.3389/fcell.2021.725071
68. Fusi G, Constantinides M, Fissoun C, et al. Senescence-Driven Inflammatory and Trophic Microenvironment Imprints Mesenchymal Stromal/Stem Cells in Osteoarthritic Patients. Biomedicines. 2023;11(7):1994. doi:10.3390/biomedicines11071994
69. Malaise O, Tachikart Y, Constantinides M, et al. Mesenchymal stem cell senescence alleviates their intrinsic and seno-suppressive paracrine properties contributing to osteoarthritis development. Aging. 2019;11(20):9128–9146. doi:10.18632/aging.102379
70. Gnani D, Crippa S, Della Volpe L, et al. An early-senescence state in aged mesenchymal stromal cells contributes to hematopoietic stem and progenitor cell clonogenic impairment through the activation of a pro-inflammatory program. Aging Cell. 2019;18(3):e12933. doi:10.1111/acel.12933
71. Fafián-Labora JA, Morente-López M, Arufe MC. Effect of aging on behaviour of mesenchymal stem cells. World J Stem Cells. 2019;11(6):337–346. doi:10.4252/wjsc.v11.i6.337
72. Chou LY, Ho CT, Hung SC. Paracrine Senescence of Mesenchymal Stromal Cells Involves Inflammatory Cytokines and the NF-κB Pathway. Cells. 2022;11(20):3324. doi:10.3390/cells11203324
73. Ye G, Li J, Yu W, et al. ALKBH5 facilitates CYP1B1 mRNA degradation via m6A demethylation to alleviate MSC senescence and osteoarthritis progression. Exp Mol Med. 2023;55(8):1743–1756. doi:10.1038/s12276-023-01059-0
74. Zhang FJ, Luo W, Lei GH. Role of HIF-1α and HIF-2α in osteoarthritis. Joint Bone Spine. 2015;82(3):144–147. doi:10.1016/j.jbspin.2014.10.003
75. Zhang XA, Kong H. Mechanism of HIFs in osteoarthritis. Front Immunol. 2023;14:1168799. doi:10.3389/fimmu.2023.1168799
76. Cowman SJ, Koh MY. Revisiting the HIF switch in the tumor and its immune microenvironment. Trends Cancer. 2022;8(1):28–42. doi:10.1016/j.trecan.2021.10.004
77. Hu S, Zhang C, Ni L, et al. Stabilization of HIF-1α alleviates osteoarthritis via enhancing mitophagy. Cell Death Dis. 2020;11(6):481. doi:10.1038/s41419-020-2680-0
78. Ryu JH, Shin Y, Huh YH, Yang S, Chun CH, Chun JS. Hypoxia-inducible factor-2α regulates Fas-mediated chondrocyte apoptosis during osteoarthritic cartilage destruction. Cell Death Differ. 2012;19(3):440–450. doi:10.1038/cdd.2011.111
79. Hu C, Li L. Preconditioning influences mesenchymal stem cell properties in vitro and in vivo. J Cell Mol Med. 2018;22(3):1428–1442. doi:10.1111/jcmm.13492
80. Yang Y, Lee EH, Yang Z. Hypoxia-Conditioned Mesenchymal Stem Cells in Tissue Regeneration Application. Tissue Eng B. 2022;28(5):966–977. doi:10.1089/ten.TEB.2021.0145
81. Samal JRK, Rangasami VK, Samanta S, Varghese OP, Oommen OP. Discrepancies on the Role of Oxygen Gradient and Culture Condition on Mesenchymal Stem Cell Fate. Adv Healthcare Mater. 2021;10(6):e2002058. doi:10.1002/adhm.202002058
82. Rosová I, Dao M, Capoccia B, Link D, Nolta JA. Hypoxic preconditioning results in increased motility and improved therapeutic potential of human mesenchymal stem cells. Stem Cells. 2008;26(8):2173–2182. doi:10.1634/stemcells.2007-1104
83. Musiał-Wysocka A, Kot M, Sułkowski M, Badyra B, Majka M. Molecular and Functional Verification of Wharton’s Jelly Mesenchymal Stem Cells (WJ-MSCs) Pluripotency. Int J Mol Sci. 2019;20(8):1807. doi:10.3390/ijms20081807
84. Mohyeldin A, Garzón-Muvdi T, Quiñones-Hinojosa A. Oxygen in stem cell biology: a critical component of the stem cell niche. Cell Stem Cell. 2010;7(2):150–161. doi:10.1016/j.stem.2010.07.007
85. Lin Q, Lee YJ, Yun Z. Differentiation arrest by hypoxia. J Biol Chem. 2006;281(41):30678–30683. doi:10.1074/jbc.C600120200
86. Adesida AB, Mulet-Sierra A, Jomha NM. Hypoxia mediated isolation and expansion enhances the chondrogenic capacity of bone marrow mesenchymal stromal cells. Stem Cell Res Ther. 2012;3(2):9. doi:10.1186/scrt100
87. Adolfsson E, Helenius G, Friberg Ö, Samano N, Frøbert O, Johansson K. Bone marrow- and adipose tissue-derived mesenchymal stem cells from donors with coronary artery disease; growth, yield, gene expression and the effect of oxygen concentration. Scand J Clin Lab Invest. 2020;80(4):318–326. doi:10.1080/00365513.2020.1741023
88. Liu H, Liu S, Li Y, et al. The role of SDF-1-CXCR4/CXCR7 axis in the therapeutic effects of hypoxia-preconditioned mesenchymal stem cells for renal ischemia/reperfusion injury. PLoS One. 2012;7(4):e34608. doi:10.1371/journal.pone.0034608
89. Xing J, Ying Y, Mao C, et al. Hypoxia induces senescence of bone marrow mesenchymal stem cells via altered gut microbiota. Nat Commun. 2018;9(1):2020. doi:10.1038/s41467-018-04453-9
90. Zhang F, Peng W, Wang T, et al. Lnc Tmem235 promotes repair of early steroid-induced osteonecrosis of the femoral head by inhibiting hypoxia-induced apoptosis of BMSCs. Exp Mol Med. 2022;54(11):1991–2006. doi:10.1038/s12276-022-00875-0
91. Sha Y, Yang L, Lv Y. ERK1/2 and Akt phosphorylation were essential for MGF E peptide regulating cell morphology and mobility but not proangiogenic capacity of BMSCs under severe hypoxia. Cell Biochem Funct. 2018;36(3):155–165. doi:10.1002/cbf.3327
92. Denu RA, Hematti P. Optimization of oxidative stress for mesenchymal stromal/stem cell engraftment, function and longevity. Free Radic Biol Med. 2021;167:193–200. doi:10.1016/j.freeradbiomed.2021.02.042
93. Waheed TO, Hahn O, Sridharan K, Mörke C, Kamp G, Peters K. Oxidative Stress Response in Adipose Tissue-Derived Mesenchymal Stem/Stromal Cells. Int J Mol Sci. 2022;23(21):13435. doi:10.3390/ijms232113435
94. Duclos M. Osteoarthritis, obesity and type 2 diabetes: the weight of waist circumference. Ann Phys Rehabil Med. 2016;59(3):157–160. doi:10.1016/j.rehab.2016.04.002
95. Khasawneh RR, Abu-El-Rub E, Almahasneh FA, et al. Addressing the impact of high glucose microenvironment on the immunosuppressive characteristics of human mesenchymal stem cells. IUBMB Life. 2023;76(5):286–295. doi:10.1002/iub.2796
96. Singh RK, Yoon DS, Mandakhbayar N, et al. Diabetic bone regeneration with nanoceria-tailored scaffolds by recapitulating cellular microenvironment: activating integrin/TGF-β co-signaling of MSCs while relieving oxidative stress. Biomaterials. 2022;288:121732. doi:10.1016/j.biomaterials.2022.121732
97. Cho H, Blatchley MR, Duh EJ, Gerecht S. Acellular and cellular approaches to improve diabetic wound healing. Adv Drug Delivery Rev. 2019;146:267–288. doi:10.1016/j.addr.2018.07.019
98. Zhu M, He X, Wang XH, et al. Complement C5a induces mesenchymal stem cell apoptosis during the progression of chronic diabetic complications. Diabetologia. 2017;60(9):1822–1833. doi:10.1007/s00125-017-4316-1
99. Sui BD, Hu CH, Zheng CX, et al. Recipient Glycemic Micro-environments Govern Therapeutic Effects of Mesenchymal Stem Cell Infusion on Osteopenia. Theranostics. 2017;7(5):1225–1244. doi:10.7150/thno.18181
100. Ren W, Chai M, Jiang M, Zhou Y, Tan W. High glucose mediates apoptosis and osteogenesis of MSCs via downregulation of AKT-Sirt1-TWIST. Mol Biol Rep. 2022;49(4):2723–2733. doi:10.1007/s11033-021-07082-6
101. Kume S, Kato S, Yamagishi S, et al. Advanced glycation end-products attenuate human mesenchymal stem cells and prevent cognate differentiation into adipose tissue, cartilage, and bone. J Bone Miner Res. 2005;20(9):1647–1658. doi:10.1359/jbmr.050514
102. Yang K, Wang XQ, He YS, et al. Advanced glycation end products induce chemokine/cytokine production via activation of p38 pathway and inhibit proliferation and migration of bone marrow mesenchymal stem cells. Cardiovasc Diabetol. 2010;9:66. doi:10.1186/1475-2840-9-66
103. Chang TC, Hsu MF, Wu KK. High glucose induces bone marrow-derived mesenchymal stem cell senescence by upregulating autophagy. PLoS One. 2015;10(5):e0126537. doi:10.1371/journal.pone.0126537
104. Zhao M, Wang S, Zuo A, et al. HIF-1α/JMJD1A signaling regulates inflammation and oxidative stress following hyperglycemia and hypoxia-induced vascular cell injury. Cell Mol Biol Lett. 2021;26(1):40. doi:10.1186/s11658-021-00283-8
105. Vono R, Jover Garcia E, Spinetti G, Madeddu P. Oxidative Stress in Mesenchymal Stem Cell Senescence: regulation by Coding and Noncoding RNAs. Antioxid Redox Signaling. 2018;29(9):864–879. doi:10.1089/ars.2017.7294
106. Castillo-Quan JI, Steinbaugh MJ, Fernández-Cárdenas LP, et al. An antisteatosis response regulated by oleic acid through lipid droplet-mediated ERAD enhancement. Sci Adv. 2023;9(1):eadc8917. doi:10.1126/sciadv.adc8917
107. Zhang Y, Wang Z, Shi B, et al. Effect of gingival mesenchymal stem cell-derived exosomes on inflammatory macrophages in a high-lipid microenvironment. Int Immunopharmacol. 2021;94:107455. doi:10.1016/j.intimp.2021.107455
108. Schaffer JE. Lipotoxicity: when tissues overeat. Curr Opin Lipidol. 2003;14(3):281–287. doi:10.1097/00041433-200306000-00008
109. Liu H, Witzigreuter L, Sathiaseelan R, et al. Obesity promotes lipid accumulation in mouse cartilage-A potential role of acetyl-CoA carboxylase (ACC) mediated chondrocyte de novo lipogenesis. J Orthopaedic Res. 2022;40(12):2771–2779. doi:10.1002/jor.25322
110. Ye X, Zhang C. Effects of Hyperlipidemia and Cardiovascular Diseases on Proliferation, Differentiation and Homing of Mesenchymal Stem Cells. Curr Stem Cell Res Ther. 2017;12(5):377–387. doi:10.2174/1574888x12666170316105805
111. Chang SF, Yeh CC, Chen PJ, Chang HI. The Impact of Lipid Types and Liposomal Formulations on Osteoblast Adiposity and Mineralization. Molecules. 2018;23(1):95. doi:10.3390/molecules23010095
112. Lin YH, Kang L, Feng WH, et al. Effects of Lipids and Lipoproteins on Mesenchymal Stem Cells Used in Cardiac Tissue Regeneration. Int J Mol Sci. 2020;21(13):4770. doi:10.3390/ijms21134770
113. Liang C, Gao S, Gao J, Xu Y, Li Q. Comparison of effects of HucMSCs, exosomes, and conditioned medium on NASH. Sci Rep. 2023;13(1):18431. doi:10.1038/s41598-023-45828-3
114. Nie YF, Shang JM, Liu DQ, et al. Apical papilla stem cell-derived exosomes regulate lipid metabolism and alleviate inflammation in the MCD-induced mouse NASH model. Biochem Pharmacol. 2024;222:116073. doi:10.1016/j.bcp.2024.116073
115. Atashi F, Modarressi A, Pepper MS. The role of reactive oxygen species in mesenchymal stem cell adipogenic and osteogenic differentiation: a review. Stem Cells Develop. 2015;24(10):1150–1163. doi:10.1089/scd.2014.0484
116. Lecka-Czernik B, Rosen CJ. Energy Excess, Glucose Utilization, and Skeletal Remodeling: new Insights. J Bone Miner Res. 2015;30(8):1356–1361. doi:10.1002/jbmr.2574
117. Liu J, Gao J, Liang Z, et al. Mesenchymal stem cells and their microenvironment. Stem Cell Res Ther. 2022;13(1):429. doi:10.1186/s13287-022-02985-y
118. Rauth S, Karmakar S, Batra SK, Ponnusamy MP. Recent advances in organoid development and applications in disease modeling. Biochim Biophys Acta Rev Cancer. 2021;1875(2):188527. doi:10.1016/j.bbcan.2021.188527
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.