")
Back to Journals » Journal of Inflammation Research » Volume 17
The Dual Role of NRF2 in Colorectal Cancer: Targeting NRF2 as a Potential Therapeutic Approach
Received 24 May 2024
Accepted for publication 22 August 2024
Published 4 September 2024 Volume 2024:17 Pages 5985—6004
DOI https://doi.org/10.2147/JIR.S479794
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Mengyun Hu,1 Lingling Yuan,1 Jie Zhu2
1School of Clinical Medicine, Chengdu University of Traditional Chinese Medicine, Chengdu, People’s Republic of China; 2Hospital of Chengdu University of Traditional Chinese Medicine, Oncology Department II, Chengdu, People’s Republic of China
Correspondence: Jie Zhu, Hospital of Chengdu University of Traditional Chinese Medicine, Oncology Department II, Chengdu, 37 Shierqiao Road, Jinniu District, Chengdu City, Sichuan Province, People’s Republic of China, Tel +86 17781808796+86, Email [email protected]
Abstract: Colorectal cancer (CRC), as the third most common bisexual cancer worldwide, requires urgent research on its underlying mechanisms and intervention methods. NRF2 is an important transcription factor involved in the regulation of redox homeostasis, protein degradation, DNA repair, and other cancer processes, playing an important role in cancer. In recent years, the complex role of NRF2 in CRC has been continuously revealed: on the one hand, it exhibits a chemopreventive effect on cancer by protecting normal cells from oxidative stress, and on the other hand, it also exhibits a protective effect on malignant cells. Therefore, this article explores the dual role of NRF2 and its related signaling pathways in CRC, including their chemical protective properties and promoting effects in the occurrence, development, metastasis, and chemotherapy resistance of CRC. In addition, this article focuses on exploring the regulation of NRF2 in CRC ferroptosis, as well as NRF2 drug modulators (activators and inhibitors) targeting CRC, including natural products, compounds, and traditional Chinese medicine formulations.
Keywords: NRF2, CRC, ferroptosis, pharmacological modulators
Graphical Abstract:

Background
NRF2 (nuclear factor (erythroid-derived 2)-like 2) is a 66-kDa transcription factor which contains a Cap’n’Collar (CNC) structure.1 It belongs to basic-ucine zipper transcription factor family, is an effective transcriptional activator that plays a core role in protecting cells from oxidative and electrophilic stress.2 The activity of NRF2 is strictly regulated by Kelch like ECH related protein 1 (KEAP1), the main negative regulator of NRF2. KEAP1 belongs to the Cul3 ubiquitin ligase family and mediates the polyubiquitination and proteasomal degradation of NRF2 protein under basic (stress free) conditions, which is the main way that KEAP1 regulates NRF2. Under cellular stress conditions, the structural integrity of the KEAP1-CUL3 ligase complex is disrupted, leading to a decrease in ubiquitination activity and an increase in cellular NRF2. Unbound NRF2 translocate to the nucleus and binds to the antioxidant response element (ARE) sequence to regulate the transcription of gene suites, including intracellular redox control, metabolic pathways, autophagy, and drug transport.3
In the past, the NRF2 signaling pathway was often considered to have the ability to act as a tumor suppressor. NRF2 enables cells to resist stresses such as carcinogens and inhibits tumor growth and progression through anti-inflammatory and antioxidant effects. It also plays an important role in protecting intestinal integrity by regulating pro-inflammatory cytokines and inducing Phase II detoxification enzymes. For example, remodelling the balance of M1/M2 subpopulations in the intestinal milieu can alleviate intestinal inflammation and prevent secondary colorectal carcinogenesis through NRF2-dependent macrophage reprogramming.4 In contrast, α-tocopherol quinone plays an important role in preventing CRC by consistently enhancing the intestinal epithelial tight junction barrier through NRF2-mediated inhibition of CLDN2 expression by STAT3 inhibition.5 However, recent studies have gradually found that the NRF2 pathway plays a role in some tumors, including colorectal cancer, in an “oncogene” like manner. Promoting cell survival, resistance to radiation, chemotherapy, and metabolic disorders, and is associated with chemotherapy resistance and poor prognosis in CRC cells. At the same time, NRF2 maintains the proliferation and invasion of cancer cells during the CRC carcinogenesis stage by metabolizing reprogramming, inhibiting cancer cell apoptosis, and enhancing the self-renewal ability of cancer stem cells. More importantly, NRF2 is widely involved in iron, lipid, amino acid, and glucose metabolism and plays an extremely important role in the regulation of ferroptosis.6 Under oxidative stress conditions, nuclear translocation of NRF2 activates downstream gene expression, leading to increased expression of antioxidant proteins, which reduces ROS and the labile iron pool (LIP) to prevent additional production of ROS to block ferroptosis. In addition, NRF2 alters iron homeostasis by increasing iron storage and its flux into and out of the cell. The intracellular iron storage protein ferritin, including ferritin heavy chain (FTH) and ferritin light chain (FTL), segregates excess free iron in protein cages, thereby limiting redox conversion of iron. Thus, NRF2 has considerable potential to regulate ferroptosis in CRC cells, which may serve as a new avenue for CRC therapy.
The Expression of NRF2 in CRC Tissues
CRC is the third most common cancer worldwide.7 Over 1.9 million new CRC cases and 930000 deaths were estimated in 2020. The burden of CRC is projected to increase to 3.2 million new cases and 1.6 million deaths by 2040 with most cases predicted to occur in high or very high HDI countries.8 The occurrence of CRC is a multi-step process that involves multiple genetic changes. This process is usually divided into three stages: initiation, promotion, and progression, and has various biological characteristics,9 such as cell transformation, genomic instability, excessive proliferation, immortalization, angiogenesis, epithelial mesenchymal transition, and metastasis. NRF2 often plays an undeniable role in these processes and the characteristics exhibited by CRC.
NRF2 is specifically elevated in CRC tissue, significantly elevated at both mRNA and protein levels, and positively correlated with tumor size, lymph nodes, distant metastasis, and smoking status.10 During the progression of adenomas to cancer, multiple genes including NRF2 and genes encoding cell cycle pathway components undergo changes.11 NRF2 gradually increases in the normal mucosal adenoma cancer direction and only shows significantly higher expression in CRC with rapid inflammatory peritumoral response, but does not show any correlation with tumor invasiveness characteristics.12 However, interestingly, research by Professor Elena V Knatko’s team has shown that NRF2 activation does not affect adenoma development in a mouse model of CRC, and NRF2 activation is unlikely to affect the early stages of CRC development.13
The expression of NRF2 exhibits significant heterogeneity based on the clinical staging and prognosis of CRC,14 making it the most promising biomarker for identifying poorly prognosis CRC. A metagene based study showed that NRF2 is a consistent and robust prognostic biomarker for all stages of CRC.15 High activity of NRF2 suggests poor prognosis in patients, especially high levels of nuclear NRF2 suggest poor prognosis.16 The cytoplasmic localization of NRF2 (cNrf2) expressed in CRC has a greater impact on tumor invasion and 5-FU resistance than nuclear localization (nNrf2), which can promote CRC invasiveness by upregulating PSMD4.17,18
In addition, KEAP1 downregulates its mRNA levels through epigenetic modification and interacts with NRF2 to increase CRC risk.19 The role of NRF2 related transcription factor heme oxygenase 1 (Hmox1) in the growth and spread of CRC cannot be ignored too. A study showed that the ratio of Hmox1/ NRF2 mRNA levels in tumor tissues of distant metastatic subjects was significantly increased compared to non-metastatic patients.20 And as a risk factor for CRC among workers, the promotion effect of red meat diet on CRC can be partially explained by initiating positive selection of precancerous cells through NRF2 dependent antioxidant response.21
The Complex Regulation of NRF2 in CRC
Not only is the abnormal expression of NRF2 itself, but the activation of NRF2 related signaling pathways is also closely related to the initiation, progression, and metastasis of CRC, and even treatment resistance. The involved pathways include classic anti-inflammatory and antioxidant pathways, as well as metabolic and ferroptosis signaling pathways that have gradually been emphasized in recent years. Therefore, here we summarize the main signal pathways regulated by NRF2 in CRC, and analyze its two-sided role in CRC.
NRF2 Mediated Chemoprevention Effects in CRC: Anti-Inflammatory and Antioxidant
Inflammation and oxidative stress play crucial roles in the occurrence and progression of various diseases, including CRC.22,23 Chronic OS can lead to oxidation of biomolecules (nucleic acids, lipids, and proteins) or activation of inflammatory signaling pathways, resulting in activation of several transcription factors or dysregulation of gene and protein expression, ultimately leading to tumor development or cancer cell survival.24
As is well known, NRF2 is a classic stress transcription factor that maintains cellular homeostasis and controls the expression of antioxidant, cell protective, and detoxifying enzymes. The balance between KEAP1 and NRF2 is crucial for maintaining cellular homeostasis, and the KEAP1/NRF2 axis is recognized as the central node for cell defense and survival pathway crosstalk.25 Under no pressure, NRF2 forms a homologous complex with KEAP1, which coexists in the cytoplasm to facilitate ubiquitination of NRF2. When oxidative or electrophilic stress exists, the oxidation of cysteine residues in KEAP1 interrupts the ubiquitination process of NRF2. Then NRF2 translocates to the nucleus, forms dimers with small Maf proteins, and binds to ARE to activate downstream gene expression, exerting antioxidant effects. Among them, NRF2 induced expression of phase II detoxifying enzymes can reduce the risk of tissue cancer and prevent early inflammation mediated tumor occurrence.26 Meanwhile, the role of the KEAP1/NRF2 signaling axis in gastrointestinal development, anti-inflammatory, and antioxidant stress has been confirmed.21 Therefore, KEAP1/NRF2 has a protective effect in the early stage of CRC, and its inhibitory carcinogenic effect may be related to activating the antioxidant defense system, regulating inflammatory mediators to weaken oxidative stress.27
The KEAP1-NRF2-ARE pathway is one of the most important defense mechanisms against oxidative and/or electrophilic stress,28 which inhibits the development of acute/chronic colitis by inhibiting inflammation and preventing oxidative stress-induced cell damage.29,30 In HCT116 cells, the activated KEAP1/NRF2 pathway can upregulate SLC7A11 expression and glutathione levels, promote lipid peroxidation and protein oxidation, leading to mitochondrial damage and ultimately iron dependent cell death.23 Meanwhile, activation of the KEAP1-NRF2-ARE pathway showed protective effects in AOM/DSS induced CRC mice.31 This indicates that the activation of the KEAP1-NRF2-ARE signaling pathway has a preventive effect on colitis related CRC. In addition, NRF2 upregulates the expression of the phase II detoxification enzyme glutathione S-transferase (including GST – α, μ), Protecting cancer progression during AOM induced CRC.31 And targeting the KEAP1/NRF2 signaling pathway can effectively reverse DMH mediated oxidative stress and DNA damage in rats.32 Specifically, the KEAP1/NRF2 signaling pathway activates major antioxidants (including enzymatic and non-enzymatic systems such as GSH, vitamin C, and E) to reduce ROS mediated cell damage and activate the transcription of phase II cell protective enzymes to upregulate intestinal barrier function, thereby playing a chemoprevention role in CRC.32–34
ARE and HO-1 are classic antioxidant factors downstream of NRF2, commonly believed to protect cells from cancer and slow down cancer development by neutralizing ROS or carcinogens.35 In vivo studies have shown that 50 mg/kg digitalis can significantly reduce the incidence, number and size of AOM-DSS induced tumors in mice, and reduce H2O2 induced oxidative stress and cell death through p38 MAPK-NRF2/ARE pathway.36 And the fermented ileal fluid rich in raspberry can drive upregulation of the cell protective NRF2-ARE pathway to reduce DNA damage in normal colon cells.37 In addition, activating the NRF2/HO-1 axis can specifically exert anti-inflammatory effects on colon tissue in an inflammatory state without affecting normal tissue,38 and demonstrate chemoprevention effects on CRC.39 In the human colorectal adenocarcinoma cell line DLD-1, the activation of NRF2-HO-1 signaling pathway induces CRC cell death through antioxidant stress, and may be related to the resistance of organobismuth (III) complex induced cell death.40 The NRF2 inhibitor Brusatol causes rapid and transient depletion of NRF2 through a KEAP1 independent post transcriptional mechanism.41 Brusatol mediated NRF2 knockdown can inhibit the expression of HO-1 induced by organobismuth (III) complex and enhance its anticancer activity.42 Aldose reductase (AR) inhibitors have been confirmed to inhibit the growth of CRC cells in cultures and xenografts in nude mice.42 Inhibition of HO-1 activity can weaken the cytotoxicity against oxidative stress and reduce the efficacy of antioxidants.43 Recent studies have shown that AR inhibitors increase the expression of NRF2 and related antioxidant enzymes in CRC cells, thereby increasing the expression of PGC-1 α, NRF1 and TFAM and reduces mitochondrial DNA damage, to regulate mitochondrial biogenesis and prevent CRC growth.44
MicroRNA-222-3p (miR-222-3p) is also a key regulatory factor for oxidative stress,45 increasing in the colon and circulating blood of CRC patients.46,47 Downregulation of miR-222-3p alleviates DSS induced oxidative damage by promoting BRG1 mediated NRF2 /HO-1 signaling in NCM460 cells, significantly alleviating oxidative stress and inflammation in the damaged colon of Ulcerative colitis (UC) and colitis-associated cancer (CAC) mice.48 In addition, lactic acid bacteria and 5-aminosalicylic acid play a protective role in ulcerative colitis models by regulating gut microbiota and the NRF2/HO-1 pathway49.
It is worth noting that during the growth and spread of cancer, there is potential crosstalk between NRF2 and the NF - κ B pathway. NF-κ B activation leads to NRF2 deficiency, resulting in increased cytokine production. Similarly, the activation of NRF2 also weakens NF- κ B and downstream signal transduction.50 Targeted activation of the NRF2-HO-1 pathway can interfere with the NF- κ B signaling pathway, reduce NF-κ B mRNA expression in colon tissue, effectively reverse the deterioration of colon mucosal ulcers, and exhibit anticancer effects in experimental CRC induced Wistar rats51,52.
NRF2 Promotes CRC Tumor Metastasis
The prognosis of CRC with or without distant metastasis varies greatly, with approximately 20% of CRC patients experiencing distant metastasis in clinical practice.53,54 Metastatic colorectal cancer is the main cause of patient mortality, with a 5-year survival rate of only about 14%.55 Research has found that, the levels of KEAP1/NRF2 and SGP proteins, as well as their T/N (tumor/normal tissue) ratio, have varying degrees of correlation in CRC subjects with or without lymph node/distant metastasis.56,57 The T/N ratio of KEAP1 protein in the NRF2/KEAP1 pathway is an important predictor of lymphatic vessel infiltration (LVI) and can serve as a negative predictor of CRC metastasis.56 Activate the KEAP1/NRF2 signaling pathway, NRF2 can directly induce the expression of miR-34a and miR-34b/c by occupying multiple ARE motifs in its promoter region, thereby inhibiting CRC metastasis.58 In addition, as a key pathogenic factor and microbial biomarker for CRC, Fusobacterium nucleatum increases CYP2J2 and 12.13-EpOME transcription by activating the TLR4/KEAP1/NRF2 axis, and then promoting EMT and metastasis of CRC59.
The activation of the NRF2/HO-1 pathway is also associated with increased invasiveness of CRC. For example, ethanol induces NRF2 and HO-1 nuclear translocation in response to oxidative stress and ER stress, endowing CRC with resistance to oxidative stress to maintain the survival of CRC cells, and inducing MMP2 and VEGF expression helps them achieve more aggressive phenotypes.60 The promotion of cancer cell proliferation and metastasis in the tumor microenvironment (TME) by M2 tumor associated macrophages (M2-TAM) is a crucial step in the progression of CRC.61 The NRF2-HO-1 axis mediated antioxidant stress in CRC selectively survives CD163+M2-TAM in TME, leading to an increase in the frequency of M2-TAM infiltration.62 Previously, Consonni FM et al’s research also showed that HO-1+CD163+M2-TAM, originating from F4/80HO-1 bone marrow precursor and dependent on NRF2 activation, promotes tumor angiogenesis and epithelial mesenchymal transition of tumor cells, and inhibits the function of CD8+T cells, leading to the formation of metastatic TME.63 In addition, salicylic acid induces AMPK and inhibits c-MYC, trans activating the NRF2/ARE/miR-34a/b/c cascade to inhibit CRC cell migration, invasion, and metastasis64.
NRF2 Participates in CRC Resistance
The resistance of cancer cells to treatment is the main cause of death in most cancer patients.65 Chemotherapy based on oxaliplatin is the standard treatment for advanced CRC patients, and oxaliplatin resistance is a significant cause of death in CRC patients.66 Research has found that,67 oxaliplatin is an activator of the Nrf2 signaling pathway, which prevents the cytotoxicity of anticancer drugs by activating the KEAP1/NRF2 antioxidant system. Similarly, inhibiting NRF2 nuclear translocation can prevent KEAP1-NRF2 activation, thereby improving the resistance of human CRC cells to oxaliplatin.68 In addition to oxaliplatin, multidrug resistance (MDR) is also a major obstacle to successful chemotherapy in CRC patients,69 and one of its common forms is caused by the activation of the ABCB1 gene and its main product P-glycoprotein. Research has found that, NRF2 is a key transcription factor that regulates efflux transporters (including P-gp), There is a positive correlation between the protein levels of NRF2 and P-gp.70 In CRC biopsy, the expression levels of NRF2 and ABCB1 (P-gp) were significantly increased at both mRNA and protein levels, while the expression levels of KEAP1 were obviously reduced in these samples.71
5-FU is the preferred chemotherapy drug for CRC, and its leading intestinal mucositis is the main limiting factor for anti-cancer treatment.72 Research has shown that CMP (Carboxymethylated pachyman, a polysaccharide extracted from traditional Chinese medicine Poria cocos) combined with 5-FU can regulate the NRF2-ARE signaling pathway to alleviate 5-FU induced colonic mucosal inflammation.73 And another study shows that, Procyanidin B2 (PB2) reduces ROS accumulation through the NRF2/ARE pathway to protect the intestinal tract of mice from radiation induced damage.74 Meanwhile, the NRF2/ARE pathway controls low-level ROS activation of β - catenin, thereby enhancing Wnt/β - catenin signaling and promoting regeneration driven by leucine rich G-protein coupled receptor 5 positive intestinal stem cells (Lgr5+ISC), achieving similar results in models of intestinal organoid injury.74 It is worth noting that, the activation of the NRF2 signaling pathway may not only be regulated by the ubiquitination of NRF2 controlled by KEAP1, but also by epigenetic modifications. Inhibiting methylation in the NRF2 promoter region can effectively activate NRF2, and the expression of NRF2 mRNA increases with a decrease in protein levels and enzyme activity of epigenetic modifying enzymes (such as DNA methyltransferase and histone deacetylase), and subsequently induces downstream antioxidant stress pathways75.
In addition, the lncRNA MIR4435-2HG levels in cisplatin resistant cell line HCT116R were significantly elevated, and knocking out MIR4435-2HG significantly restored sensitivity to cisplatin.76 Meanwhile, the mRNA levels of NRF2 and HO-1 are inhibited with the knockout of MIR4435-2HG, indicating that lncRNA MIR4435-2HG mediates cisplatin resistance through NRF2/HO-1.76 Similarly, inhibiting the expression of NRF2/HO-1 pathway and pathway related biomarkers can reverse 5-FU resistance in colon cancer cells.77
Other Regulation of NRF2 in CRC
In addition to participating in CRC resistance, metastasis, and early cell protective effects, NRF2 can also intervene in CRC progression by regulating metabolism, autophagy, and DNA damage. The ROS released by mitochondria activates NRF2, which affects CRC glycolysis, oxidative phosphorylation, mitochondrial biogenesis, and mitochondrial autophagy through the KEAP1/NRF2 pathway, and affects tumor metastasis.78 ADSL expression prominently increases in CRC, indicating involvement in DNA synthesis DNA repair and upregulation of cell cycle genes accelerate cell cycle progression and significantly increase proliferation and migration of CRC cell lines. KEAP1-NRF2 is activated when ADSL is overexpressed, which is beneficial for the survival and proliferation of ROS accumulating cells, as well as DNA damage and tumorigenesis.79 In addition, low levels of ITLN1 have been shown to be associated with obesity in the development of CRC and are independent prognostic factors for CRC.80 ITLN1 inhibits tumor derived IL-17D mediated tumor neovascularization, bone marrow-derived EPC recruitment, and MDSC production and transportation through the KEAP1/NRF2/ROS/IL-17D signaling cascade dependent on PI3K/AKT/GSK3 ß,81 thereby benefiting CRC metastasis and immune suppression.
The Pleckstrin homology (PH) domain rich leucine repeat protein phosphatase 2 (PHLPP2) is a key regulator of cellular homeostasis and plays a tumor suppressive role in various human cancers.82 In CRC, PHLPP2 can inhibit the stemness of CRC cells by inhibiting the NRF2-ARE signaling pathway and increasing ROS levels.83 In addition, sodium butyrate dose-dependently inhibits the growth of CRC HCT116 cells, at least partially due to inhibition of the NRF2 signaling pathway.84 Sodium butyrate has the potential to demethylate the promoter of KEAP1 gene, leading to an increase in KEAP1 gene expression, thereby blocking NRF2-ARE signal transduction and NRF2 target gene expression, weakening the biological function of NRF2, and inducing CRC cell apoptosis by activating caspase to inhibit Bcl-xL protein.85
The sustained activation of NRF2, which is beneficial to cancer cells, is known as “NRF2 addiction”86 and is typically beneficial for CRC progression. Therefore, intervention targeting CRC cells with NRF2 addiction may become a new therapeutic target. Research shows that, L-selenocysteine can reduce NRF2 and autophagy pathway protein expression, selectively attacking NRF2 addicted cancer cells and ultimately leading to cell death.87 It is worth mentioning that activating the KEAP1/NRF2 pathway to induce protective antioxidant stress response can effectively respond to the accumulation of ROS caused by radiotherapy and improve the effectiveness of radiotherapy.88
The Regulation of NRF2 in CRC Ferroptosis
The Relationship Between NRF2 and CRC Ferroptosis: As a Major Negative Regulatory Factor
Ferroptosis is a newly discovered form of iron and reactive oxygen species dependent regulation of cell death, involving iron metabolism disorders and ROS accumulation in the plasma membrane. Ferroptosis is mainly induced by intracellular iron accumulation and lipid peroxidation. Excessive iron generates ROS through Fenton reaction to promote cell ferroptosis, which is accompanied by GSH depletion.
NRF2 is the key to cellular antioxidant response, widely involved in iron, lipid, amino acid, glucose metabolism, etc., and plays an extremely important role in the regulation of ferroptosis.6 Physiologically, the nuclear translocation of NRF2 activates the expression of genes under oxidative stress conditions, resulting in increased expression of antioxidant proteins, thereby reducing ROS and the LIP, to block the occurrence of ferroptosis (Figure 1). Therefore, NRF2 is the main negative regulator of ferroptosis, leading to abnormal inhibition of ferroptosis in tumors. Specifically, the downstream gene HO-1 of NRF2 has a dual role in ferroptosis, increasing LIP while producing antioxidants. It is worth noting that recent studies have gradually revealed the non-negligible role of ferroptosis in tumor suppression (Figure 1). Therefore, inducing ferroptosis in tumor cells by regulating NRF2 and related pathways has become a promising new method for treating cancer, including CRC.
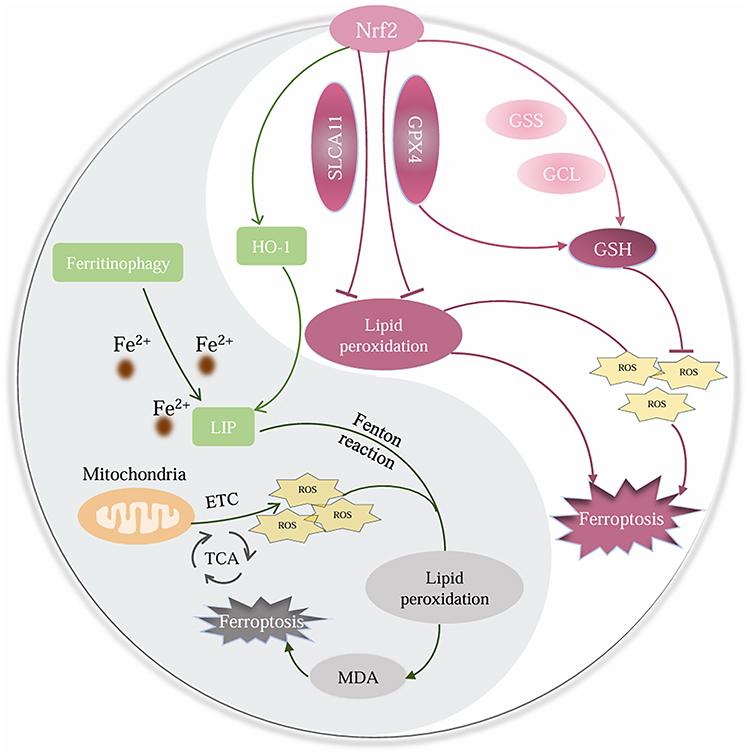 |
Figure 1 The Classic Mechanism of Ferroptosis and the Role of NRF2 in Regulating CRC Ferroptosis. |
NRF2 Regulates CRC Ferroptosis
Glutathione peroxidase 4 (GPX4) is one of the most important antioxidant enzymes, regulated by NRF2. GPX4, as a key regulatory factor of ferroptosis, has attracted considerable attention in fields such as cancer, cardiovascular, and neuroscience in the past decade.89 In preclinical studies, the regulatory mechanism of GPX4 level/activity has shown great potential for the treatment of ferroptosis related diseases. For example, upregulation of NRF2/GPX4 signaling pathway can affect the onset of pre-eclampsia by inhibiting ferroptosis,90 and can save the cognitive dysfunction of diabetes encephalopathy.91 While inhibiting NRF2/GPX4 pathway to induce cell ferroptosis can accelerate female postmenopausal atherosclerosis,92 and can induce lung cancer cells and liver cells ferroptosis.93,94 In addition, inhibiting the NRF2/GPX4 pathway can enhance the sensitivity of multiple drug-resistant tumors to chemotherapy drugs and promote their ferroptosis.95–97 However, the significant impact of the NRF2/GPX4 signaling pathway in CRC has not been emphasized and summarized in previous articles. Based on this, we will summarize the role of the NRF2/GPX4 pathway in CRC ferroptosis.
Forkhead box transcription factor A2 (FOXA2) is a transcription factor that plays a crucial role in embryonic development, metabolic homeostasis and tumor cell proliferation. FOXA2 expression was significantly up-regulated in tumor samples of CRC patients, and was positively correlated with NRF2/GPX4 gene expression.98 FOXA2 up-regulated indicating that the survival rate of CRC patients was poor.98 In terms of mechanism, FOXA2 promotes CRC progression by inhibiting NRF2/GPX4 ferroptosis signaling pathway. Conversely, depletion of FOXA2 can weaken the activation of NRF2 pathway and reduce the level of GPX4 in CRC cells, thereby inducing ferroptosis.98 Studies have shown that,99 the expression of transferrin increased and the expression of GPX4 decreased in CRC cells treated with RSL3 (ferroptosis inducer). On the contrary, GPX4 overexpression attenuated RSL3 induced FCD. This suggests that decreasing GPX4 by inhibiting NRF2 may be the core factor of RSL3 induced ferroptosis in CRC cells.
The resistance of CRC to oxaliplatin is predictable, and the potential mechanism of its occurrence may involve inhibition of ferroptosis. An experiment targeting acquired oxaliplatin resistant CRC cells (HCT116 Or) as well as congenital resistant (H716) CRC cells revealed that inhibition of cell ferroptosis by the KIF20A/NUAK1/PP1 β/ GPX4 pathway may be the basis of oxaliplatin resistance in CRC.100 And inducing ferroptosis can significantly reverse oxaliplatin resistance in CRC cells.100 It is worth noting that although apoptosis remains the main process of cell death in sensitized cancer cells in Caco-2 cell lines, GPX4/siRNA induced ferroptosis still kills cancer cells through alternative pathways to overcome oxaliplatin resistance.101
In addition to the NRF2/GPX4 signaling pathway regulating ferroptosis in CRC cells, other signaling pathways are also widely involved in the ferroptosis mechanism of CRC. Research has shown that activation of the PERK-NRF2-HO-1 signaling pathway mediated by ER stress can induce ferroptosis in CRC cells, accompanied by a decrease in GSH levels and an increase in lipid peroxidation.102 Under ER stress conditions, endoplasmic reticulum stress-related kinase (PERK) directly phosphorylates NRF2. Subsequently, phosphorylated NRF2 translocates to the nucleus and trans activates its effector HO-1 expression, leading to an increase in unstable iron pools and promoting lipid peroxidation, resulting in early ferroptosis in HCT116 cells.102 Furthermore, cetuximab enhances RSL3 induced ferroptosis by activating p38 MAPK and inhibiting NRF2/HO-1 axis.99 It can be seen that inducing ferroptosis in CRC cells through the NRF2-HO-1 signaling pathway may expand the efficacy and scope of chemotherapy drugs. At the same time, ferroptosis can selectively target invasive cancer stem cells and has the potential to enhance the efficacy of immunotherapy, overcoming resistance to immunotherapy.103
Sodium butyrate (NaB) can significantly increase RSL3 induced cell ferroptosis, and its sensitivity to ferroptosis is determined by FFAR2 -mTOR signaling transduction.104 Specifically, NaB mediates the downregulation of SLC7A11 and GPX4 in a cAMP-PKA dependent manner through the FFAR2-AKT-NRF2 axis and FFAR2-mTORC1 axis, respectively, inducing lipid ROS production and promoting ferroptosis.104 And ginsenoside Rh3 triggers CRC cell pyroptosis and ferroptosis through the Stat3/p53/NRF2 axis, with almost no damage to normal cells.105 While uridine cytidine kinase like-1 (uckl1), which is highly expressed in a variety of cancers, has become a classical suppressor of CRC cell ferroptosis by stabilizing NRF2 and promoting SLC7A11 expression, playing an atypical role in inhibiting CRC cell ferroptosis106.
In summary inducing ferroptosis in tumor cells has been widely studied as a promising new method for combating drug-resistant cancer. Inducing CRC ferroptosis through the NRF2 related signaling pathway may be beneficial for cancer patients by inhibiting oxaliplatin resistance, promoting tumor cell ferroptosis, and enhancing immunotherapy efficacy.
Targeted NRF2 in CRC Treatment and Prevention
The bidirectional regulatory role of NRF2 and related signaling pathways in CRC has been elaborated in detail in the previous section. On the one hand, the NRF2 signaling pathway can play a chemoprevention role in the occurrence of CRC, especially CAC, through anti-inflammatory and antioxidant mechanisms. Meanwhile, chronic toxin exposure, protein-protein interactions, epigenetic modification factors, transcription/translation regulatory factors, post-translational modifications, and mutations can also induce NRF2 in CRC for a long time and promote cancer occurrence, progression, metastasis, and chemotherapy resistance. More importantly, the activation and inhibition of the NRF2 signaling pathway may be beneficial for CRC in different microenvironments. Therefore, despite the considerable complexity and difficulty of targeting NRF2 in cancer, finding ways to manipulate each individual mode of NRF2 activation remains a key strategy for preventing or slowing down tumorigenesis, decreasing NRF2 addiction, cancer invasiveness and ultimately improving patient prognosis. Here, we summaries the NRF2 inhibitors and activators involved in CRC in recent years (including natural products, compounds, and some Chinese medicinal preparations) to serve as a reference for further research (Tables 1 and 2).Preclinical studies to intervene in CRC evolution by modulating NRF2 activity have been quite abundant, as we have summarized in detail in the table Meanwhile, relevant clinical studies are gradually emerging. For example, the glutaminase inhibitor CB-839 in combination with 5-FU increased reactive oxygen species and induced nuclear translocation of NRF2, which in turn up-regulated the mRNA expression of uridine phosphorylase I, facilitated the conversion of 5-FU to its active compounds, and thus enhanced the inhibitory effect on thymidylate synthase and induced tumor regression in PIK3CA-mutant CRCs in multiple xenograft models.107 And the combination of CB-839 and capecitabine, a prodrug of 5-FU, was demonstrated to be well tolerated with antitumor activity at biologically active doses in a subsequent Phase I clinical trial.107 A prospective randomised controlled study experimentally showed that L-myostatin significantly elevated NRF2 levels (38.7%) and reduced the levels/activity of NF-κB (27%) and TNF-α (36.6%), which was neuroprotective against oxaliplatin-induced peripheral neuropathy in colorectal cancer patients by targeting the NRF2 and NF-κB pathways.108 In contrast, the NEDD8-activating enzyme inhibitor Pevonedistat (TAK-924/MLN4924) exhibits clinical anti-tumor effects by non- specifically increasing NRF2 protein accumulation in patients with advanced solid tumours, including colorectal cancer.109,110 In addition, a novel synthetic triterpenoid and antioxidant inflammatory modulator, Bardoxolone methyl, effectively induced NRF2 and inhibited NF-κB and JAK/STAT signaling pathway, and was able to activate NQO1, a key target of NRF2, in tumour tissues at the mRNA level, and clinical anticancer activity was observed in patients with solid tumors.111
 |
Table 1 NRF2 Activators |
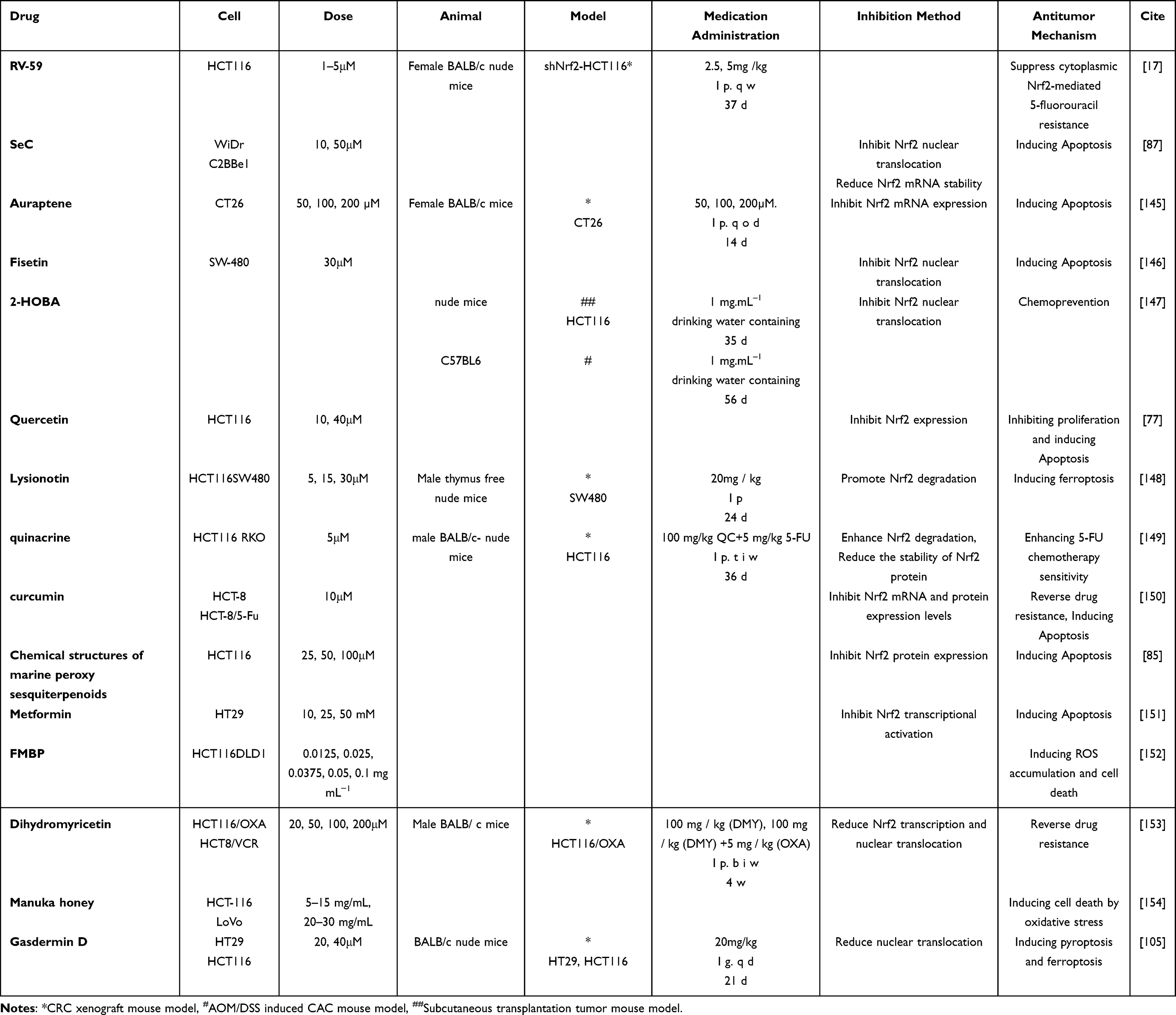 |
Table 2 NRF2 Inhibitor |
Summary and Future Directions
In conclusion, we summarized the opposite effects of Nrf2 and its related signaling pathways in the development and progression of CRC, which may be related to different stages of CRC and its heterogeneous tumor microenvironment. During the progression from adenoma to carcinoma, mutations occur in genes involved in nine driver signaling pathways, including the NRF2 signaling pathway.11 In an immunohistochemical examination of 93 clinical tissues (65 carcinomas with corresponding surgical margins and 28 adenomas), it was found that NRF2 was progressively elevated in the direction of what appeared to be a normal mucosa-adenoma-carcinoma and showed significantly higher expression only in CRC with a rapid inflammatory peritumoral response, which may reveal a role for NRF2 in CRC carcinogenesis.12 Furthermore, there was a significant difference between NRF2 expression and host immune response in adenoma specimens, with mean NRF2 expression correlating with high levels of dysplasia, indicating a poor response to treatment.12 These studies demonstrate the complexity of the role of NRF2 in CRC, which is a double-edged sword. On the one hand, NRF2 resists malignant transformation of cells by maintaining the “redox balance” at normal levels, demonstrating a chemoprevention role. On the other hand, after malignant transformation, high NRF2 expression induces the transcription of antioxidants, giving them a pro-survival phenotype and ultimately the formation of chemo-resistant tumors.
The main body of chemoprevention effects of NRF2 appears in the inflammation-cancer sequence progression of CAC. Chronic inflammation strongly contributes to the onset and progression of CAC by persistently stimulating colonic epithelial cells (IECs) and inducing heterotypic proliferation through multiple mechanisms, including the induction of genetic changes, oxidative stress-driven DNA damage, aberrant immune responses, and disruption of intestinal flora.155 The NRF2 pathway is involved in the maintenance of intestinal development and normal function, the development of UC, and UC-associated intestinal fibrosis and oncogenic effects, and mice deficient in NRF2 will have increased susceptibility to dextrose sodium sulfate-induced colitis and CRC.156 In addition, inhibition of the signaling pathway of NRF2, a key transcription factor associated with cellular antioxidant responses and a central regulator in the maintenance of intracellular redox homeostasis, will disrupt the integrity of the intestinal barrier and induce colon tumorigenesis.48 Therefore, activation of the NRF2 signaling pathway to alleviate oxidative stress in IEC can prevent and ameliorate UC and CAC occurrence. For example, in the context of oxidative damage in CAC mice, the activities of the antioxidant enzymes GSH-Px and SOD are significantly reduced, and ROS and MDA levels are significantly elevated in colonic tissues.48 Increasing NRF2 expression increases the activity of the antioxidant enzyme SOD, decreases the level of MDA, and inhibits the levels of inflammatory markers NF-κB, TNF-α, IL-1β, and MPO in CAC mice, which has a preventive effect on the occurrence of CAC.48 Activation of some classical pathways promotes the oncogenic effects of UC-CAC, and functional crosstalk between both NRF2 and NF-к B was found.157 By activating NRF2-mediated expression of phase II detoxification enzymes and reducing NF-κ B-associated inflammation could improve the intestinal environment, mucosal barrier, colonic and crypt disruption, and reduce ulceration and microbial translocation, which could reduce CAC occurrence.158 In addition, regulation of NRRF2 modulates various genes involved in cellular redox, protein degradation, DNA repair, xenobiotic metabolism, and apoptosis, again contributing to the prevention of CAC.158 All these evidences reveal the promising application of regulating NRRF2 activity in the prevention of CAC.
In addition to the classic chemo-preventive effect (anti-inflammatory and antioxidant), as well as the key role of Nrf2 in CRC progression, metastasis and drug resistance, we especially pay attention to the great potential of Nrf2 in CRC cell “ erroptosis”. Notably, mismatch repair deficient and microsatellite instability high (D-MMR/MSI-H) represents a unique biomarker-defined group of cancers with higher tumor neoantigen loads and denser immune cell infiltration compared to mismatch repair proficient and microsatellite stable (p MMR/MSS) tumours, and is considered a positive immune checkpoint inhibitor (ICI) efficacy predictor. However, D-MMR/MSI-H-type CRCs account for only about 15% of all types of CRCs, and most MSS-type CRCs respond poorly to ICI therapy.159 Studies have shown that the MSI score of CRC is negatively correlated with the NRF2 expression program,160 revealing the negative role of NRF2 in anti-tumor immunity. TMB values correlate significantly with NRF2/KEAP1 mutations in nine cancer subtypes including CRC,161 and compared with unmutated cancers, cancers with mutations in PIK3CA, a gene associated with the NRF2 pathway, had a higher CRC had higher TMB.162 Furthermore, a subpopulation of NRF2 mutations in cancers is associated with poor prognosis but with increased PD-L1 expression, which may be beneficial for ICI therapy.161 However, although NRF2 mutations are associated with TMB-H, a negative correlation between NRF2 mutations and immune reactivity profile scores, negative ICI predictors, and CD8T cell infiltration may impair anti-tumor immune responses in MSS-type CRC.160,162 Therefore, modulating NRF2 expression in CRC and remodelling the CRC immune microenvironment may be an effective means to intervene in CRC progression.
In summary, for different CRC stages and tumor microenvironment, selective use of Nrf2 inhibitor activators will greatly improve the current situation and difficulties of CRC treatment. However, a large number of existing studies are still based on basic research at the cellular level and animal level, and further rigorous clinical research is still needed. We hope that through this review, researchers can see the broad prospects of NRF2 and its related signaling pathways in the application of CRC, and encourage more researchers to conduct more in-depth research to provide more solid and powerful evidence for the clinical application of NRF2 activators or inhibitors.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Jaramillo MC, Zhang DD. The emerging role of the Nrf2-Keap1 signaling pathway in cancer. Genes Dev. 2013;27(20):2179–2191.
2. Kobayashi EH, Suzuki T, Funayama R, et al. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat Commun. 2016;7:11624.
3. He F, Ru X, Wen T. NRF2, a transcription factor for stress response and beyond. Int J Mol Sci. 2020;21(13):1.
4. Lu C, Xue L, Luo K, et al. Colon-accumulated gold nanoclusters alleviate intestinal inflammation and prevent secondary colorectal carcinogenesis via nrf2-dependent macrophage reprogramming. ACS Nano. 2023;17(18):18421–18432.
5. Ganapathy AS, Saha K, Wang A, et al. Alpha-tocopherylquinone differentially modulates claudins to enhance intestinal epithelial tight junction barrier via AhR and Nrf2 pathways. Cell Rep. 2023;42(7):112705.
6. Yan R, Lin B, Jin W, Tang L, Hu S, Cai R. NRF2, a Superstar of Ferroptosis. Antioxidants. 2023;12(9):1.
7. Duan N, Hu X, Qiu H, et al. Targeting the E2F1/Rb/HDAC1 axis with the small molecule HR488B effectively inhibits colorectal cancer growth. Cell Death & Disease. 2023;14(12):801.
8. Morgan E, Arnold M, Gini A, et al. Global burden of colorectal cancer in 2020 and 2040: incidence and mortality estimates from GLOBOCAN. Gut. 2023;72(2):338–344.
9. Hanahan D. Hallmarks of Cancer: new Dimensions. Cancer Discov. 2022;12(1):31–46.
10. Sadeghi MR, Jeddi F, Soozangar N, et al. Nrf2/P-glycoprotein axis is associated with clinicopathological characteristics in colorectal cancer. Biomed Pharm. 2018;104:458–464.
11. Hu Y, Chen Y, Guo H, et al. Molecular alterations in circulating cell-free DNA in Patients with Colorectal Adenoma or Carcinoma. Cancer Manag Res. 2020;12:5159–5167.
12. El-Deek HEM, Ahmed AM, Mohammed RAA. Aberration of Nrf2‑Bach1 pathway in colorectal carcinoma; role in carcinogenesis and tumor progression. Annals Diagnostic Path. 2019;38:138–144.
13. Knatko EV, Castro C, Higgins M, et al. Nrf2 activation does not affect adenoma development in a mouse model of colorectal cancer. Communi Biology. 2021;4(1):1081.
14. Ji L, Wei Y, Jiang T, Wang S. Correlation of Nrf2, NQO1, MRP1, cmyc and p53 in colorectal cancer and their relationships to clinicopathologic features and survival. Intern J Clini Experim Path. 2014;7(3):1124–1131.
15. O’Cathail SM, C-H W, Lewis A, Holmes C, Hawkins MA, Maughan T. NRF2 metagene signature is a novel prognostic biomarker in colorectal cancer. Cancer Genetics. 2020:248–249.
16. Torrente L, Maan G, Oumkaltoum Rezig A, et al. High NRF2 Levels Correlate with Poor Prognosis in Colorectal Cancer Patients and with Sensitivity to the Kinase Inhibitor AT9283 In Vitro. Biomolecules. 2020;10(10).
17. Shen C-J, Lin P-L, Lin H-C, Cheng Y-W, Huang H-S, Lee H. RV-59 suppresses cytoplasmic Nrf2-mediated 5-fluorouracil resistance and tumor growth in colorectal cancer. Ame J Cancer Res. 2019;9(12):2789–2796.
18. Lin P-L, Chang JT, D-W W, Huang -C-C, Lee H. Cytoplasmic localization of Nrf2 promotes colorectal cancer with more aggressive tumors via upregulation of PSMD4. Free Radi Biol Med. 2016;95:121–132.
19. Gao L, Yuan F, Che G, et al. Epigenetic modifications but not genetic polymorphisms regulate KEAP1 expression in colorectal cancer. J Cell bioch. 2019;120(8):12311–12320.
20. Chang L-C, Fan C-W, Tseng W-K, et al. The Ratio of Hmox1/Nrf2 mRNA Level in the Tumor Tissue Is a Predictor of Distant Metastasis in Colorectal Cancer. Disease Mark. 2016;2016:8143465.
21. Surya R, Héliès-Toussaint C, Martin OC, et al. Red meat and colorectal cancer: nrf2-dependent antioxidant response contributes to the resistance of preneoplastic colon cells to fecal water of hemoglobin- and beef-fed rats. Carcinogenesis. 2016;37(6):635–645.
22. Basak D, Uddin MN, Hancock J. The Role of Oxidative Stress and Its Counteractive Utility in Colorectal Cancer (CRC). Cancers. 2020;12(11).
23. Yang S, Li W, Bai X, et al. Ginseng-derived nanoparticles alleviate inflammatory bowel disease via the TLR4/MAPK and p62/Nrf2/Keap1 pathways. J Nanob. 2024;22(1):48.
24. Bardelčíková A, Šoltys J, Mojžiš J. Oxidative Stress, Inflammation and Colorectal Cancer: an Overview. Antioxidants. 2023;12(4).
25. Bian L, Nguyen VT, Tamaoki J, et al. Genetic hyperactivation of Nrf2 causes larval lethality in Keap1a and Keap1b-double-knockout zebrafish. Redox Biology. 2023;62:102673.
26. Osburn WO, Karim B, Dolan PM, et al. Increased colonic inflammatory injury and formation of aberrant crypt foci in Nrf2-deficient mice upon dextran sulfate treatment. Intern J Can. 2007;121(9):1883–1891.
27. Piotrowska M, Swierczynski M, Fichna J, Piechota-Polanczyk A. The Nrf2 in the pathophysiology of the intestine: molecular mechanisms and therapeutic implications for inflammatory bowel diseases. Pharmacological Research. 2021;163:105243.
28. Shahcheraghi SH, Salemi F, Peirovi N, et al. Nrf2 Regulation by Curcumin: molecular Aspects for Therapeutic Prospects. Molecules. 2021;27(1).
29. M-C L, Ji J-A, Jiang Y-L, et al. An inhibitor of the Keap1-Nrf2 protein-protein interaction protects NCM460 colonic cells and alleviates experimental colitis. Scientific Reports. 2016;6:26585.
30. Wang X, Saud SM, Zhang X, Li W, Hua B. Protective effect of Shaoyao Decoction against colorectal cancer via the Keap1-Nrf2-ARE signaling pathway. Journal of Ethnopharmacology. 2019;241:111981.
31. Pandurangan AK, Ananda Sadagopan SK, Dharmalingam P, Ganapasam S. Luteolin, a bioflavonoid inhibits Azoxymethane-induced colorectal cancer through activation of Nrf2 signaling. Toxic Mech Meth. 2014;24(1):13–20.
32. Darband SG, Sadighparvar S, Yousefi B, et al. Quercetin attenuated oxidative DNA damage through NRF2 signaling pathway in rats with DMH induced colon carcinogenesis. Life Sciences. 2020;253:117584.
33. Wu Y, Wang D, Yang X, Fu C, Zou L, Zhang J. Traditional Chinese medicine Gegen Qinlian decoction ameliorates irinotecan chemotherapy-induced gut toxicity in mice. Biomed Pharmacother. 2019;109:2252–2261.
34. Pandurangan AK, Saadatdoust Z, Esa NM, Hamzah H, Ismail A. Dietary cocoa protects against colitis-associated cancer by activating the Nrf2/Keap1 pathway. BioFactors. 2015;41(1).
35. Zhao P, Wei Y, Sun G, et al. Fetuin-A alleviates neuroinflammation against traumatic brain injury-induced microglial necroptosis by regulating Nrf-2/HO-1 pathway. J Neuroinflammation. 2022;19(1):269.
36. Yang Y, Cai X, Yang J, et al. Chemoprevention of dietary digitoflavone on colitis-associated colon tumorigenesis through inducing Nrf2 signaling pathway and inhibition of inflammation. Mol Cancer. 2014;13:48.
37. Dobani S, Latimer C, McDougall GJ, et al. Ex vivo fecal fermentation of human ileal fluid collected after raspberry consumption modifies (poly)phenolics and modulates genoprotective effects in colonic epithelial cells. Redox Biology. 2021;40:101862.
38. Kang S, Kim W, Jeong S, et al. Oxidized 5-aminosalicylic acid activates Nrf2-HO-1 pathway by covalently binding to Keap1: implication in anti-inflammatory actions of 5-aminosalicylic acid. Free Radical Biology & Medicine. 2017;108:715–724.
39. Bi W, C-N H, X-X L, et al. Ginnalin A from Kujin tea (Acer tataricum subsp. ginnala) exhibits a colorectal cancer chemoprevention effect via activation of the Nrf2/HO-1 signaling pathway. Food & function. 2018;9(5):2809–2819.
40. Iuchi K, Tasaki Y, Shirai S, Hisatomi H. Upregulation of nuclear factor (erythroid-derived 2)-like 2 protein level in the human colorectal adenocarcinoma cell line DLD-1 by a heterocyclic organobismuth(III) compound: effect of organobismuth(III) compound on NRF2 signaling. Biomed Pharmacothera. 2020;125:109928.
41. Cai SJ, Liu Y, Han S, Yang C. Brusatol, an NRF2 inhibitor for future cancer therapeutic. Cell & bioscience. 2019;9:45.
42. Sonowal H, Pal PB, Wen -J-J, Awasthi S, Ramana KV, Srivastava SK. Aldose reductase inhibitor increases doxorubicin-sensitivity of colon cancer cells and decreases cardiotoxicity. Scientific Reports. 2017;7(1):3182.
43. Kwon DH, Cha H-J, Lee H, et al. Protective Effect of Glutathione against Oxidative Stress-induced Cytotoxicity in RAW 264.7 Macrophages through Activating the Nuclear Factor Erythroid 2-Related Factor-2/Heme Oxygenase-1 Pathway. Antioxidants. 2019;8(4).
44. Shukla K, Sonowal H, Saxena A, Ramana KV, Srivastava SK. Aldose reductase inhibitor, fidarestat regulates mitochondrial biogenesis via Nrf2/HO-1/AMPK pathway in colon cancer cells. Cancer Letters. 2017;411:57–63.
45. Wang J, Luo X, Cai S, Sun J, Wang S, Wei X. Blocking HOTAIR protects human chondrocytes against IL-1β-induced cell apoptosis, ECM degradation, inflammatory response and oxidative stress via regulating miR-222-3p/ADAM10 axis. Inter Immunopharm. 2021;98:107903.
46. Liu S, Sun X, Wang M, et al. A microRNA 221- and 222-mediated feedback loop maintains constitutive activation of NFκB and STAT3 in colorectal cancer cells. Gastroenterology. 2014;147(4).
47. Gasparello J, Papi C, Allegretti M, et al. A Distinctive microRNA (miRNA) Signature in the Blood of Colorectal Cancer (CRC) Patients at Surgery. Cancers. 2020;12(9).
48. Wang X-J, Zhang D, Yang Y-T, et al. Suppression of microRNA-222-3p ameliorates ulcerative colitis and colitis-associated colorectal cancer to protect against oxidative stress via targeting BRG1 to activate Nrf2/HO-1 signaling pathway. Frontiers in Immunology. 2023;14:1089809.
49. El-Baz AM, Khodir AE, Adel El-Sokkary MM, Shata A. The protective effect of Lactobacillus versus 5-aminosalicylic acid in ulcerative colitis model by modulation of gut microbiota and Nrf2/Ho-1 pathway. Life Sciences. 2020;256:117927.
50. Wardyn JD, Ponsford AH, Sanderson CM. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem Soc Trans. 2015;43(4):621–626.
51. Rehman MU, Rashid S, Arafah A, et al. Piperine Regulates Nrf-2/Keap-1 Signalling and Exhibits Anticancer Effect in Experimental Colon Carcinogenesis in Wistar Rats. Biology. 2020;9(9).
52. Abo El-Fadl HM I, Mohamed MFA. Targeting endoplasmic reticulum stress, Nrf-2/HO-1, and NF-κB by myristicin and its role in attenuation of ulcerative colitis in rats. Life Sciences. 2022;311(Pt B):121187.
53. Zhao W, Dai S, Yue L, et al. Emerging mechanisms progress of colorectal cancer liver metastasis. Front Endoc. 2022;13:1081585.
54. Zhou H, Liu Z, Wang Y, et al. Colorectal liver metastasis: molecular mechanism and interventional therapy. Signal Transduct Target Ther. 2022;7(1):70.
55. Shin AE, Giancotti FG, Rustgi AK. Metastatic colorectal cancer: mechanisms and emerging therapeutics. Trends Pharmacol Sci. 2023;44(4):222–236.
56. Chang L-C, Fan C-W, Tseng W-K, Chen J-R, Hua -C-C. The tumour/normal tissue ratio of Keap1 protein is a predictor for lymphovascular invasion in colorectal cancer: a correlation study between the Nrf2 and KRas pathways. Biomarkers. 2022;27(7):701–707.
57. Chang L-C, Fan C-W, Tseng W-K, Hua -C-C. The level of S-glutathionylated protein is a predictor for metastasis in colorectal cancer and correlated with those of Nrf2/Keap1 pathway. Biomarker. 2021;26(8):780–787.
58. Liu C, Rokavec M, Huang Z, Hermeking H. Curcumin activates a ROS/KEAP1/NRF2/miR-34a/b/c cascade to suppress colorectal cancer metastasis. Cell Death Differ. 2023;30(7):1771–1785.
59. Wang N, Fang J-Y. Fusobacterium nucleatum, a key pathogenic factor and microbial biomarker for colorectal cancer. Trends Microbiol. 2023;31(2):159–172.
60. Cernigliaro C, D’Anneo A, Carlisi D, et al. Ethanol-Mediated Stress Promotes Autophagic Survival and Aggressiveness of Colon Cancer Cells via Activation of Nrf2/HO-1 Pathway. Cancers. 2019;11(4).
61. He X, Chen H, Zhong X, et al. BST2 induced macrophage M2 polarization to promote the progression of colorectal cancer. Inter J Biol Scien. 2023;19(1):331–345.
62. Ito M, Mimura K, Nakajima S, et al. M2 tumor-associated macrophages resist to oxidative stress through heme oxygenase-1 in the colorectal cancer tumor microenvironment. Cancer Immu Immuno. 2023;72(7):2233–2244.
63. Consonni FM, Bleve A, Totaro MG, et al. Heme catabolism by tumor-associated macrophages controls metastasis formation. Nat Immunol. 2021;22(5):595–606.
64. Liu C, Rokavec M, Huang Z, Hermeking H. Salicylate induces AMPK and inhibits c-MYC to activate a NRF2/ARE/miR-34a/b/c cascade resulting in suppression of colorectal cancer metastasis. Cell Death & Disease. 2023;14(10):707.
65. Debaugnies M, Rodríguez-Acebes S, Blondeau J, et al. RHOJ controls EMT-associated resistance to chemotherapy. Nature. 2023;(7955):168–175.
66. Chen G, Gong T, Wang Z, et al. Colorectal cancer organoid models uncover oxaliplatin-resistant mechanisms at single cell resolution. Cell Oncol. 2022;45(6):1155–1167.
67. Wang XJ, Li Y, Luo L, et al. Oxaliplatin activates the Keap1/Nrf2 antioxidant system conferring protection against the cytotoxicity of anticancer drugs. Free Radical biol Med. 2014;70:68–77.
68. Tanaka S, Hosokawa M, Tatsumi A, Asaumi S, Imai R, Ogawara K-I. Improvement of resistance to oxaliplatin by vorinostat in human colorectal cancer cells through inhibition of Nrf2 nuclear translocation. Biochem Biophys Rese Communi. 2022;607.
69. Chang SN, Kang SC. Decursinol Angelate Inhibits Glutamate Dehydrogenase 1 Activity and Induces Intrinsic Apoptosis in MDR-CRC Cells. Cancers. 2023;15(14).
70. Shchulkin AV, Abalenikhina YV, Kosmachevskaya OV, Topunov AF, Yakusheva EN. Regulation of P-Glycoprotein during Oxidative Stress. Antioxidants. 2024;13(2).
71. Mostafazadeh M, Kahroba H, Haiaty S, et al. In vitro exosomal transfer of Nrf2 led to the oxaliplatin resistance in human colorectal cancer LS174T cells. Cell biochem Func. 2022;40(4):391–402.
72. Xia J, He S, Dai Q, et al. Atorvastatin calcium alleviates 5-fluorouracil-induced intestinal damage by inhibiting cellular senescence and significantly enhances its antitumor efficacy. Intern Immunopha. 2023;121:110465.
73. Wang C, Yang S, Gao L, Wang L, Cao L. Carboxymethyl pachyman (CMP) reduces intestinal mucositis and regulates the intestinal microflora in 5-fluorouracil-treated CT26 tumour-bearing mice. Food & Function. 2018;9(5):2695–2704.
74. Zhu X, Tian X, Yang M, et al. Procyanidin B2 Promotes Intestinal Injury Repair and Attenuates Colitis-Associated Tumorigenesis via Suppression of Oxidative Stress in Mice. Antioxidants & Redox Signaling. 2021;35(2):75–92.
75. Zuo Q, Wu R, Xiao X, et al. The dietary flavone luteolin epigenetically activates the Nrf2 pathway and blocks cell transformation in human colorectal cancer HCT116 cells. Journal of Cellular biochemistry. 2018;119(11):9573–9582.
76. Luo P, Wu S, Ji K, et al. LncRNA MIR4435-2HG mediates cisplatin resistance in HCT116 cells by regulating Nrf2 and HO-1. PLoS One. 2020;15(11):e0223035.
77. Tang Z, Wang L, Chen Y, et al. Quercetin reverses 5-fluorouracil resistance in colon cancer cells by modulating the NRF2/HO-1 pathway. European J Histoch. 2023;67(3).
78. Chang L-C, Fan C-W, Tseng W-K, Hua -C-C. Associations between the Nrf2/Keap1 pathway and mitochondrial functions in colorectal cancer are affected by metastasis. Canc Biomark. 2020;27(2):163–171.
79. Taha-Mehlitz S, Bianco G, Coto-Llerena M, et al. Adenylosuccinate lyase is oncogenic in colorectal cancer by causing mitochondrial dysfunction and independent activation of NRF2 and mTOR-MYC-axis. Theranostics. 2021;11(9):4011–4029.
80. Kawashima K, Maeda K, Saigo C, Kito Y, Yoshida K, Takeuchi T. Adiponectin and Intelectin-1: important Adipokine Players in Obesity-Related Colorectal Carcinogenesis. Int J Mol Sci. 2017;18(4).
81. Chen L, Jin X-H, Luo J, et al. ITLN1 inhibits tumor neovascularization and myeloid derived suppressor cells accumulation in colorectal carcinoma. Oncogene. 2021;40(40):5925–5937.
82. Jin H, Ma J, Xu J, et al. Oncogenic role of MIR516A in human bladder cancer was mediated by its attenuating PHLPP2 expression and BECN1-dependent autophagy. Autophagy. 2021;17(4):840–854.
83. Yongfu X, He Z, Xujian H, et al. PLHPP2 inhibits the stemness of colorectal cancer by inactivating the Nrf2 signaling pathway. Journal of Cancer. 2022;13(4):1313–1323.
84. Wang L, Shannar AAF, Wu R, et al. Butyrate Drives Metabolic Rewiring and Epigenetic Reprogramming in Human Colon Cancer Cells. Mole Nutrit Food Rese. 2022;66(12):e2200028.
85. Taira J, Miyazato H, Ueda K. Marine Peroxy Sesquiterpenoids Induce Apoptosis by Modulation of Nrf2-ARE Signaling in HCT116 Colon Cancer Cells. Marine Drugs. 2018;16(10).
86. Schmidlin CJ, Shakya A, Dodson M, Chapman E, Zhang DD. The intricacies of NRF2 regulation in cancer. Semin Cancer Biol. 2021;76:110–119.
87. Hsu W-L, Wang C-M, Yao C-L, et al. Blockage of Nrf2 and autophagy by L-selenocystine induces selective death in Nrf2-addicted colorectal cancer cells through p62-Keap-1-Nrf2 axis. Cell Death & Disease. 2022;13(12):1060.
88. Bader S, Wilmers J, Pelzer M, Jendrossek V, Rudner J. Activation of anti-oxidant Keap1/Nrf2 pathway modulates efficacy of dihydroartemisinin-based monotherapy and combinatory therapy with ionizing radiation. Free Radical Biology & Medicine. 2021;168:44–54.
89. Xie Y, Kang R, Klionsky DJ, Tang D. GPX4 in cell death, autophagy, and disease. Autophagy. 2023;19(10):2621–2638.
90. Liao T, Xu X, Ye X, Yan J. DJ-1 upregulates the Nrf2/GPX4 signal pathway to inhibit trophoblast ferroptosis in the pathogenesis of preeclampsia. Scientific Reports. 2022;12(1):2934.
91. Shi Y-S, Chen J-C, Lin L, et al. Dendrobine rescues cognitive dysfunction in diabetic encephalopathy by inhibiting ferroptosis via activating Nrf2/GPX4 axis. Phytomedicine. 2023;119:154993.
92. Lv Y, Zhang S, Weng X, et al. Estrogen deficiency accelerates postmenopausal atherosclerosis by inducing endothelial cell ferroptosis through inhibiting NRF2/GPX4 pathway. FASEB. 2023;37(6):e22992.
93. Cai S, Ding Z, Liu X, Zeng J. Trabectedin induces ferroptosis via regulation of HIF-1α/IRP1/TFR1 and Keap1/Nrf2/GPX4 axis in non-small cell lung cancer cells. Chem Biol Interact. 2023;369:110262.
94. Wang X, Zhu W, Xing M, Zhu H, Chen E, Zhou J. Matrine disrupts Nrf2/GPX4 antioxidant system and promotes hepatocyte ferroptosis. Chem Biol Interact. 2023;384:110713.
95. C-S M, Q-M L, Zhang K-R, et al. NRF2-GPX4/SOD2 axis imparts resistance to EGFR-tyrosine kinase inhibitors in non-small-cell lung cancer cells. Acta Pharmacol Sin. 2021;42(4):613–623.
96. Huang W, Wen F, Yang P, Li Y, Li Q, Shu P. Yi-qi-hua-yu-jie-du decoction induces ferroptosis in cisplatin-resistant gastric cancer via the AKT/GSK3β/NRF2/GPX4 axis. Phytomedicine. 2024;123:155220.
97. Zhang Z, Hu Q, Ye S, Xiang L. Inhibition of the PIN1-NRF2/GPX4 axis imparts sensitivity to cisplatin in cervical cancer cells. Acta biochimica et biophysica Sinica. 2022;54(9):1325–1335.
98. Liu X, Yan C, Chang C, et al. FOXA2 Suppression by TRIM36 Exerts Anti-Tumor Role in Colorectal Cancer Via Inducing NRF2/GPX4-Regulated Ferroptosis. Advan Sci. 2023;10(35):e2304521.
99. Yang J, Mo J, Dai J, et al. Cetuximab promotes RSL3-induced ferroptosis by suppressing the Nrf2/HO-1 signalling pathway in KRAS mutant colorectal cancer. Cell Death & Disease. 2021;12(11):1079.
100. Yang C, Zhang Y, Lin S, Liu Y, Li W. Correction for: suppressing the KIF20A/NUAK1/Nrf2/GPX4 signaling pathway induces ferroptosis and enhances the sensitivity of colorectal cancer to oxaliplatin. Aging. 2021;13(14):19077.
101. Golbashirzadeh M, Heidari HR, Aghamolayi AA, Fattahi Y, Talebi M, Khosroushahi AY. In vitro siRNA-mediated GPX4 and AKT1 silencing in oxaliplatin resistance cancer cells induces ferroptosis and apoptosis. Medical Oncol. 2023;40(10):279.
102. Wei R, Zhao Y, Wang J, et al. Tagitinin C induces ferroptosis through PERK-Nrf2-HO-1 signaling pathway in colorectal cancer cells. Int J Bio Sci. 2021;17(11):2703–2717.
103. Elgendy SM, Alyammahi SK, Alhamad DW, et al. Ferroptosis: an emerging approach for targeting cancer stem cells and drug resistance. Crit Rev Oncol Hematol. 2020;155:103095.
104. Wang G, Qin S, Chen L, et al. Butyrate dictates ferroptosis sensitivity through FFAR2-mTOR signaling. Cell Death & Disease. 2023;14(4):292.
105. Wu Y, Pi D, Zhou S, et al. Ginsenoside Rh3 induces pyroptosis and ferroptosis through the Stat3/p53/NRF2 axis in colorectal cancer cells. Acta biochimica et biophysica Sinica. 2023;55(4):587–600.
106. Wu W, Zhao Y, Qin B, et al. Non-canonical role of UCKL1 on ferroptosis defence in colorectal cancer. EBioMedicine. 2023;93:104650.
107. Zhao Y, Feng X, Chen Y, et al. 5-Fluorouracil Enhances the Antitumor Activity of the Glutaminase Inhibitor CB-839 against PIK3CA-Mutant Colorectal Cancers. Cancer Research. 2020;80(21):4815–4827.
108. Yehia R, Saleh S, El Abhar H, Saad AS, Schaalan M. L-Carnosine protects against Oxaliplatin-induced peripheral neuropathy in colorectal cancer patients: a perspective on targeting Nrf-2 and NF-κB pathways. Toxicol Appl Pharmacol. 2019;365:41–50.
109. Andérica-Romero AC, Hernández-Damián J, Vázquez-Cervantes GI, Torres I, González-Herrera IG, Pedraza-Chaverri J. The MLN4924 inhibitor exerts a neuroprotective effect against oxidative stress injury via Nrf2 protein accumulation. Redox Biology. 2016;8:341–347.
110. Sarantopoulos J, Shapiro GI, Cohen RB, et al. Phase I Study of the Investigational NEDD8-Activating Enzyme Inhibitor Pevonedistat (TAK-924/MLN4924) in Patients with Advanced Solid Tumors. Clin Cancer Res. 2016;22(4):847–857.
111. Hong DS, Kurzrock R, Supko JG, et al. A phase I first-in-human trial of bardoxolone methyl in patients with advanced solid tumors and lymphomas. Clin Cancer Res. 2012;18(12):3396–3406.
112. Wang F, Dezfouli AB, Khosravi M, et al. Cannabidiol-induced crosstalk of apoptosis and macroautophagy in colorectal cancer cells involves p53 and Hsp70. Cell Death Discovery. 2023;9(1):286.
113. Ren X, Zhang G, Ling X, et al. Allyl-isothiocyanate against colorectal cancer via the mutual dependent regulation of p21 and Nrf2. European Journal of Pharmacology. 2023;957:176016.
114. Liang J, Yang C, Li P, et al. Astragaloside IV inhibits AOM/DSS-induced colitis-associated tumorigenesis via activation of PPARγ signaling in mice. Phytomedicine. 2023;121:155116.
115. Deng Y, Huang X, Chen X, et al. Chemopreventive Effects of Polysaccharides and Flavonoids from Okra Flowers in Azomethane/Dextran Sulfate Sodium-Induced Murine Colitis-Associated Cancer. Nutrients. 2023;15(22).
116. Chang C-H, Lien Y-T, Lin W-S, Nagabhushanam K, C-T H, Pan M-H. Protective Effects of Piceatannol on DNA Damage in Benzo[a]pyrene-Induced Human Colon Epithelial Cells. J Agr Food Chem. 2023;71(19):7370–7381.
117. Baptistella MM, Assunção RRS, Sales de Oliveira C, et al. A synthetic resveratrol-curcumin hybrid derivative exhibits chemopreventive effects on colon pre-neoplastic lesions by targeting Wnt/β-catenin signaling, anti-inflammatory and antioxidant pathways. J Pharm. 2023.
118. Jeong S, Farag AK, Yun HK, et al. AF8c, a Multi-Kinase Inhibitor Induces Apoptosis by Activating DR5/Nrf2 via ROS in Colorectal Cancer Cells. Cancers. 2022;14(13).
119. Khan MI, Karima G, Khan MZ, Shin JH, Kim JD. Therapeutic Effects of Saponins for the Prevention and Treatment of Cancer by Ameliorating Inflammation and Angiogenesis and Inducing Antioxidant and Apoptotic Effects in Human Cells. Int J Mol Sci. 2022;23(18).
120. Dong M, Liu H, Cao T, et al. Huoxiang Zhengqi alleviates azoxymethane/dextran sulfate sodium-induced colitis-associated cancer by regulating Nrf2/NF-κB/NLRP3 signaling. Frontiers in Pharmacology. 2022;13:1002269.
121. Batjargal A, Solek P, Kukula-Koch W, et al. Gurgem-7 toxicity assessment: regulation of cell survival or death by traditional Mongolian prescription. Ecotoxicol Environm Safe. 2022;239:113660.
122. Serrya MS, El-Sheakh AR, Makled MN. Evaluation of the therapeutic effects of mycophenolate mofetil targeting Nrf-2 and NLRP3 inflammasome in acetic acid induced ulcerative colitis in rats. Life Sciences. 2021;271:119154.
123. Papierska K, Krajka-Kuźniak V, Paluszczak J, et al. Lichen-Derived Depsides and Depsidones Modulate the Nrf2, NF-κB and STAT3 Signaling Pathways in Colorectal Cancer Cells. Molecules. 2021;26(16).
124. Ishak NIM, Mohamed S, Madzuki IN, Mustapha NM, Esa NM. Limonin modulated immune and inflammatory responses to suppress colorectal adenocarcinoma in mice model. Naunyn-Schmiedeberg’s Archives of Pharm. 2021;394(9):1907–1915.
125. Wang X, Saud SM, Wang F, et al. Protective effect of ShaoYao decoction on colitis-associated colorectal cancer by inducing Nrf2 signaling pathway. Journal of Ethnopharmacology. 2020;252:112600.
126. Lee D-Y, Yun S-M, Song M-Y, Jung K, Kim E-H. Cyanidin Chloride Induces Apoptosis by Inhibiting NF-κB Signaling through Activation of Nrf2 in Colorectal Cancer Cells. Antioxidants. 2020;9(4).
127. Hao Q, Wang M, Sun N-X, et al. Sulforaphane suppresses carcinogenesis of colorectal cancer through the ERK/Nrf2‑UDP glucuronosyltransferase 1A metabolic axis activation. Oncology Reports. 2020;43(4):1067–1080.
128. Chaparala A, Tashkandi H, Chumanevich AA, et al. Molecules from American Ginseng Suppress Colitis through Nuclear Factor Erythroid-2-Related Factor 2. Nutrients. 2020;12(6).
129. Lee K-M, Shin JM, Chun J, Song K, Nho CW, Kim YS. Igalan induces detoxifying enzymes mediated by the Nrf2 pathway in HepG2 cells. Journal of Biochemical and Molecular Toxicology. 2019;33(5):e22297.
130. Zheng Z, Chen Y, Huang J, Deng H, Tang X, Wang XJ. Mkp-1 is required for chemopreventive activity of butylated hydroxyanisole and resveratrol against colitis-associated colon tumorigenesis. Food Chemical Toxicol. 2019;127:72–80.
131. Oronsky B, Scribner C, Aggarwal R, Cabrales P. RRx-001 protects normal tissues but not tumors via Nrf2 induction and Bcl-2 inhibition. J Cancer Res Clinical Onco. 2019;145(8):2045–2050.
132. Kang KA, Piao MJ, Hyun YJ, et al. Luteolin promotes apoptotic cell death via upregulation of Nrf2 expression by DNA demethylase and the interaction of Nrf2 with p53 in human colon cancer cells. Experim Molec Med. 2019;51(4):1.
133. Jang CH, Moon N, Oh J, Kim J-S. Luteolin shifts oxaliplatin-induced cell cycle arrest at G₀/G₁ to Apoptosis in HCT116 human colorectal carcinoma cells. Nutrients. 2019;11(4):1.
134. Fang R, Wu R, Zuo Q, et al. Sophora flavescens Containing-QYJD formula activates nrf2 anti-oxidant response, blocks cellular transformation and protects against DSS-induced colitis in mouse model. American J Chinese Med. 2018;2018:1.
135. Wu X, Song M, Gao Z, et al. Nobiletin and its colonic metabolites suppress colitis-associated colon carcinogenesis by down-regulating iNOS, inducing antioxidative enzymes and arresting cell cycle progression. J Nutrit Bioch. 2017;42:17–25.
136. Onuma W, Asai D, Tomono S, et al. Anticarcinogenic effects of dried citrus peel in colon carcinogenesis due to inhibition of oxidative stress. Nutrition and Cancer. 2017;69(6):855–861.
137. Noma N, Fujii G, Miyamoto S, et al. Impact of acetazolamide, a carbonic anhydrase inhibitor, on the development of intestinal polyps in min mice. Int J Mol Sci. 2017;18(4):1.
138. Trivedi PP, Jena GB, Tikoo KB, Kumar V. Melatonin modulated autophagy and Nrf2 signaling pathways in mice with colitis-associated colon carcinogenesis. Molecular Carcinogenesis. 2016;55(3):255–267.
139. Sekar V, Anandasadagopan SK, Ganapasam S. Genistein regulates tumor microenvironment and exhibits anticancer effect in dimethyl hydrazine-induced experimental colon carcinogenesis. Bio Factors. 2016;42(6):623–637.
140. Havermann S, Chovolou Y, Humpf H-U, Wätjen W. Modulation of the Nrf2 signalling pathway in Hct116 colon carcinoma cells by baicalein and its methylated derivative negletein. Pharmac Biol. 2016;54(9):1491–1502.
141. Tan BL, Norhaizan ME, Huynh K, Yeap SK, Hazilawati H, Roselina K. Brewers’ rice modulates oxidative stress in azoxymethane-mediated colon carcinogenesis in rats. World J Gastroenterol. 2015;21(29):8826–8835.
142. Manigandan K, Manimaran D, Jayaraj RL, Elangovan N, Dhivya V, Kaphle A. Taxifolin curbs NF-κB-mediated Wnt/β-catenin signaling via up-regulating Nrf2 pathway in experimental colon carcinogenesis. Biochimie. 2015;119:103–112.
143. Yao J, Zhao L, Zhao Q, et al. NF-κB and Nrf2 signaling pathways contribute to wogonin-mediated inhibition of inflammation-associated colorectal carcinogenesis. Cell Death Disease. 2014;5(6):e1283.
144. Huber S, Valente S, Chaimbault P, Schohn H. Evaluation of ∆2-pioglitazone, an analogue of pioglitazone, on colon cancer cell survival: evidence of drug treatment association with autophagy and activation of the Nrf2/Keap1 pathway. Internat J Oncol. 2014;45(1):426–438.
145. Ebrahimi S, Soukhtanloo M, Mostafavi-Pour Z. Anti-tumor effects of Auraptene through induction of apoptosis and oxidative stress in a mouse model of colorectal cancer. Tissue & Cell. 2023;81:102004.
146. Pandey A, Trigun SK. Fisetin induces apoptosis in colorectal cancer cells by suppressing autophagy and down-regulating nuclear factor erythroid 2-related factor 2 (Nrf2). J Cell Biochem. 2023;124(9):1289–1308.
147. Gobert AP, Asim M, Smith TM, et al. Electrophilic reactive aldehydes as a therapeutic target in colorectal cancer prevention and treatment. Oncogene. 2023;42(20):1685–1691.
148. Gao Z, Jiang J, Hou L, Ji F. lysionotin induces ferroptosis to suppress development of colorectal cancer via promoting Nrf2 degradation. Oxid Med Cell Long. 2022;2022:1366957.
149. Kim HG, Kim CW, Lee DH, Lee J-S, Oh E-T, Park HJ. Quinacrine-mediated inhibition of Nrf2 reverses hypoxia-induced 5-fluorouracil resistance in colorectal cancer. Int J Mol Sci. 2019;20(18):1.
150. Zhang C, L-J H, H-Z Y, et al. Nrf2 is a key factor in the reversal effect of curcumin on multidrug resistance in the HCT‑8/5‑Fu human colorectal cancer cell line. Molecular Medicine Reports. 2018;18(6):5409–5416.
151. Sena P, Mancini S, Benincasa M, Mariani F, Palumbo C, Roncucci L. Metformin induces apoptosis and alters cellular responses to oxidative stress in ht29 colon cancer cells: preliminary findings. Int J Mol Sci. 2018;19(5):1.
152. Shan S, Shi J, Li Z, et al. Targeted anti-colon cancer activities of a millet bran-derived peroxidase were mediated by elevated ROS generation. Food & Function. 2015;6(7):2331–2338.
153. Wang Z, Sun X, Feng Y, et al. Dihydromyricetin reverses MRP2-induced multidrug resistance by preventing NF-κB-Nrf2 signaling in colorectal cancer cell. Phytomedicine. 2021;82:153414.
154. Afrin S, Giampieri F, Forbes-Hernández TY, et al. Manuka honey synergistically enhances the chemopreventive effect of 5-fluorouracil on human colon cancer cells by inducing oxidative stress and apoptosis, altering metabolic phenotypes and suppressing metastasis ability. Free Radical Biol Med. 2018;126:41–54.
155. Fantini MC, Guadagni I. From inflammation to colitis-associated colorectal cancer in inflammatory bowel disease: pathogenesis and impact of current therapies. Dig Liver Dis. 2021;53(5):558–565.
156. Peng S, Shen L, Yu X, et al. The role of Nrf2 in the pathogenesis and treatment of ulcerative colitis. Frontiers in Immunology. 2023;14:1200111.
157. Prakash AN, Prasad N, Puppala ER, et al. Loganic acid protects against ulcerative colitis by inhibiting TLR4/NF-κB mediated inflammation and activating the SIRT1/Nrf2 anti-oxidant responses in-vitro and in-vivo. Int Immunopharmacol. 2023;122:110585.
158. Laurindo LF, de Maio MC, Minniti G, et al. Effects of medicinal plants and phytochemicals in Nrf2 pathways during inflammatory bowel diseases and related colorectal cancer: a comprehensive review. Metabolites. 2023;13(2):1.
159. Li J, Wu C, Hu H, et al. Remodeling of the immune and stromal cell compartment by PD-1 blockade in mismatch repair-deficient colorectal cancer. Cancer Cell. 2023;41(6):1.
160. Wang J, Xiu J, Farrell A, et al. Mutational analysis of microsatellite-stable gastrointestinal cancer with high tumour mutational burden: a retrospective cohort study. Lancet Oncol. 2023;24(2):151–161.
161. Xu X, Yang Y, Liu X, et al. NFE2L2/KEAP1 mutations correlate with higher tumor mutational burden Value/PD-L1 expression and potentiate improved clinical outcome with immunotherapy. Oncologist. 2020;25(6):e955–e963.
162. Li N, Zhan X. Machine learning identifies pan-cancer landscape of nrf2 oxidative stress response pathway-related genes. Oxidative Medic Cel Lon. 2022;2022:8450087.
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.