")
Back to Journals » Journal of Inflammation Research » Volume 18
The Modulation of Neuroimmune Responses in Peripheral Inflammation
Authors Wang J, Ji D, Dai N, Qin C, Gong M, Fu B
Received 9 April 2025
Accepted for publication 1 July 2025
Published 10 July 2025 Volume 2025:18 Pages 9015—9030
DOI https://doi.org/10.2147/JIR.S533106
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Jing Wang,1 Dandan Ji,2 Ninan Dai,1 Chang Qin,1 Mingyang Gong,1 Bao Fu1
1Department of Critical Care Medicine, Affiliated Hospital of Zunyi Medical University, Zunyi, Guizhou, People’s Republic of China; 2Department of Intensive Care Unit, Zhejiang Province People’s Hospital Bijie Hospital, Bijie, Guizhou, People’s Republic of China
Correspondence: Bao Fu, Department of Critical Care Medicine, Affiliated Hospital of Zunyi Medical University, Dalian Road149, Zunyi, Guizhou, 563000, People’s Republic of China, Tel +86 851 2860 8514, Email [email protected]
Abstract: The brain, as the “commander-in-chief” of the human body, is known to pick up the peripheral situation and dictate orders to the periphery on time, and the immune system makes no exception. Inflammation is a defensive reaction of the body, which can be beneficial, however, unrestricted inflammation can result in life-threatening injuries and multi-organ dysfunction. The intricate interaction between the nervous and the immune system would prevent inflammation from spreading indefinitely. The onset of life-threatening bursts of inflammation may indicate neurological dysregulation, at which point additional interventions are necessary to establish a new balance. However, these interventions must be predicated on an understanding of the neuroimmune communication. Consequently, we provide a comprehensive pathway that illustrates how the central nervous system detects peripheral inflammatory signals that are transmitted by nerves or related substances and subsequently regulates peripheral inflammation through the autonomic nervous system and neuroendocrine system, intending to discover new methods of treating peripheral inflammation by intervening the nervous system.
Keywords: central nervous system, peripheral inflammation, vagus nerve, neuroimmune
Introduction
Peripheral inflammation is a kind of defense mechanism, and the body uses it to combat the invasion and attack of external harmful substances. In the inflammatory reflex, whether it is pro-inflammatory or anti-inflammatory, the secretion of cytokines is essential.1 Because the pro-inflammatory cytokine storm may lead to systemic inflammatory response syndrome (SIRS), which brings more serious injuries than the initial stimulus, it is extremely important to accurately assess the extent of its increase.2 Compensatory reactive anti-inflammatory syndrome (CRAS) may be due to the excessive production of anti-inflammatory cytokines, and anti-inflammatory cytokines will suppress the immune system and increase the risk of infection.3 Previous studies have proved that the degree of inflammation can be controlled through neuroimmune interaction, to achieve local damage control and prevent further damage.1,4 The onset of peripheral inflammation can be regarded as the result of the synergistic action of the nervous system and the immune system. This process can be summarized into three main steps: the perception and transmission of peripheral inflammatory signals, the synthesis and coordination of the received inflammatory signals by the central nervous system (CNS), and the transmission of the synthesized information from the CNS to the periphery to regulate the immune system. These steps involve extremely complex molecular mechanisms and procedures. Of course, there is local neuroimmune regulation in the periphery.5
Both animal studies and clinical studies have shown that peripheral inflammation can be alleviated by interfering with the nervous system.6–8 However, due to the current lack of sufficient understanding of the underlying mechanisms, our ability to target the nervous system for intervention in the treatment of peripheral inflammation has been greatly limited. This review aims to describe the progress of research into the mechanisms of bidirectional regulation between the CNS and peripheral inflammation, with the goal of achieving an in-depth understanding based on current studies. It explores therapeutic approaches to enhance the effectiveness of CNS interventions in the clinical management of peripheral inflammation and improve patient prognoses.
Inflammatory Signals: Perception and Input
Nerve Conduction
A wide range of substances in the peripheral circulation can transmit peripheral inflammatory signals to the CNS in multiple ways, thereby triggering a variety of responses in the CNS. These substances are produced during inflammation, including a wide range of inflammatory and anti-inflammatory cytokines, chemokines, and other related molecules. In addition to serving as agents in the transmission of signals, they also induce a variety of co-occurring changes in illness behaviors (as detailed below). This inflammatory signaling is partially mediated by vagal sensory fibers. Cytokines, pathogen products, and other inflammatory molecules secreted by peripheral immune cells can act on the tumor necrosis factor (TNF) receptors, IL-1β receptors, Fc receptors, Toll-like receptors, or pattern recognition receptors (PRR) on the vagus nerve afferent nerves to transmit these signals to the CNS.9–11 These signals are specific because different clusters of nerves within the afferent vagus nerve are responsible for encoding information about identified cytokines.12 This explains how the brain generates distinct responses to different inflammatory cytokines. Elucidating the specific markers of neural clusters that respond to different inflammatory mediators could enable the precise neuroimmune control of peripheral inflammation and reduce the side effects of neurological interventions.
The vagus nerve afferent nerve is activated by peripheral inflammatory cytokines and projects to the nucleus tractus solitarius (NTS) in the medulla oblongata of the brainstem.13 There, it continues to stimulate the release of glutamate by excitatory glutamatergic neurons in the NTS, thereby transmitting inflammatory information to the CNS.14,15 In addition to the excitotoxic effect of glutamate on CNS neurons, glutamate can also stimulate the NR2B receptor on mast cells, resulting in the secretion of lots of neuropeptides and inflammatory mediators.16 These mediators directly cause neuronal destruction or indirectly through the activation of glial cells. Subsequently, impaired neurons and activated glial cells will release a diverse array of neuropeptides and inflammatory mediators, which will continue to activate the brain mast cells. This persistent malignant positive feedback process results in a consistent rise in inflammatory mediators, which in turn triggers neuroinflammation.17,18 It seems that activating vagal afferent nerves due to peripheral inflammation increases the levels of glutamate in the CNS, leading to a certain degree of neurotoxicity.
Nociceptors found on the free nerve endings of primary sensory neurons in the dorsal root ganglia play a role in inflammatory signaling. For example, TRPA1 and TRPV1, which are part of the transient receptor potential ion channel family, are present throughout the peripheral and CNS.19 When these receptors are activated, they have a specific anti-inflammatory effect.20,21 Furthermore, their simultaneous activation results in a synergistic effect.22
BBB Destruction and Peripheral Immune Cell Infiltration
Aside from relaying inflammatory signals to the CNS through the stimulation of peripheral sensory nerves, it is probable that there exist alternative routes for the CNS to respond and regulate peripheral inflammation.23 The BBB is a crucial physiological structure that maintains homeostasis in the intracerebral environment by separating the CNS from the peripheral circulation. It is composed of vascular endothelial cells, pericytes, basement membrane, and astrocytes.24 The effects of peripheral inflammation on the CNS require investigation of this crucial physiological structure because these structures cooperate to block the penetration of peripheral hazards, and any abnormality in one of them can disrupt CNS homeostasis (Figure 1).
 |
Figure 1 Transmission of peripheral inflammatory signals via the BBB or CSF. The red stars indicate directions that merit further exploration. Abbreviations: NDS, nanoparticle delivery systems; EVs, extracellular vesicles; CSF, cerebrospinal fluid. |
Under normal physiological conditions, the peripheral circulatory system exchanges substances with the brain bilaterally by BBB.25 And leukocytes are typically found within the blood vessels and do not enter healthy brain tissue.26 However, under pathological conditions where there is peripheral inflammation, the damage to the endothelial cell caused by various inflammatory cytokines, the changes in tight junctions between endothelial cells, the activation of astrocytes and microglia, the enhancement of oxidative stress, and the alterations of transport pathways and receptors can disrupt the BBB integrity and increase the transportation of certain hazardous substances.27,28 Thereby peripheral immune cells and cytokines can cross the BBB and infiltrate into the brain.26,28,29 The BBB demonstrates a heightened sensitivity to the presence of TNFα in the peripheral blood, leading to increased permeability of the BBB and the migration of monocytes.30 The infiltration of peripheral blood monocytes into the hippocampus is a means of transmitting peripheral inflammatory signals to the CNS through cell-cell contact between monocytes and hippocampal tissue, and this process involves the release of pro-inflammatory cytokine by monocytes.31 Furthermore, the infiltrating monocytes interact with glial cells and neurons, resulting in neuroinflammation and hyperreactivity.31,32 The neuroinflammation that was induced by peripheral LPS injection was inhibited following the depletion of monocytes.33
Research by the team of Vanessa Y. Ruiz, employing single-cell RNA sequencing analysis of in vitro matured human CD14+CD16+ monocytes, revealed a distinct subset exhibiting a gene expression signature associated with CNS inflammatory diseases. This subset demonstrated a preferential capacity for migration across an in vitro human BBB model.34 Concurrently, researchers explore cell-mediated nanoparticle delivery systems conjugated to monocytes for brain tumour therapy.35,36 Consequently, beyond pharmacological strategies reducing monocyte infiltration to mitigate neuroinflammation, the intrinsic capacity of specific monocyte subsets to penetrate the BBB and infiltrate brain tissue could be leveraged. Combining this with nanoparticle delivery systems offers the potential for more precise therapeutic interventions.
Additionally, peripheral T and B cell infiltration into the brain via the compromised BBB not only causes neuronal demyelination but also participates in central immune surveillance.37,38 Peripheral inflammatory signals take time to cross the BBB and reach the CNS, but perivascular cells (PDGFRβ+ cells), key BBB and neurovascular unit components, can detect them within two hours of infection. By releasing the chemokine CCL2, these cells rapidly boost glutamatergic neuron firing and excitatory synaptic transmission in various brain regions for an initial response.39
Other Pathways
There are other additional paths for the transfer of messages at the periphery-central interface, but the two pathways mentioned above are the primary routes for the transmission of peripheral inflammatory messages to CNS. The choroid plexus is a component of the blood-cerebrospinal fluid barrier and is responsible for the production of cerebrospinal fluid (CSF). CSF also functions as a medium for peripheral inflammatory signal transmission, as evidenced by the observation that, in the stimulation of peripheral inflammation, choroid plexus-secreted extracellular vesicles or proinflammatory extracellular vesicles originating from other tissue transmit inflammatory signals to the brain via CSF.40,41 These vesicles can be taken up by astrocytes and microglia, resulting in the inhibition of miRNA target inhibition and the upregulation of inflammatory genes.41,42 However, further exploration is required to elucidate the specific mechanism by which peripheral inflammation leads to the secretion of these extracellular vesicles. Moreover, the identification of a clear biomarker for these extracellular vesicles could pave the way for the development of novel treatment targets for peripheral inflammation (Figure 1). Periventricular organs, which are regions of the brain that lack the BBB, such as the area postrema (AP), are composed of ependymal cells and non-neuronal Cox2+ cells that can respond to inflammatory cytokines in the peripheral circulation and secrete prostaglandins to transmit inflammatory signals to neurons.43,44
Overall, although this signal transmission can potentially cause some adverse effects on CNS, the next response of the brain is dependent on the central perception of peripheral inflammation signals.
Brain Responses to Peripheral Inflammation
Microglia Activation
Microglia are a type of immune cell that are distributed throughout the whole brain and conduct complex and close two-way communication with other brain tissues, such as neurons and astroglia. Consequently, microglia are not only responsible for immune function, but also exhibit heightened sensitivity to alterations in the brain microenvironment, thereby contributing to the maintenance of brain stability.45,46 Microglia have a role in the pathological processes of many different CNS disorders. As the disease progresses, its phenotype expression, morphology, and function are extremely dynamic, and this is also the case in pathological scenarios involving peripheral inflammation.47,48 The inflammatory information transmitted peripherally can stimulate microglia through the influence on synaptic connections and neuronal networks in the brain, or the secretion of inflammatory substances and other cytokines.49,50
The activation of microglia in the brain is induced by peripheral inflammation in both the early and late stages of the disease, and this effect can persist for an extended period.51–53 The activation of microglia induces the disruption of the BBB, the raised expression of pro-inflammatory cytokines in the brain, the reduction of hippocampal synaptic plasticity, the onset of oxidative stress, and neuronal death in the brain.54–56 The reduction of cell death and the amelioration of the impairment of synaptic plasticity in the hippocampus were achieved by inhibiting microglia activation, which also resulted in the reversal of the decline in the expression of anti-apoptotic proteins and the reduction of central and peripheral inflammatory cytokines.57,58 To address this abnormal activation of microglia upon peripheral inflammation, it is crucial to promptly restore him to a state of equilibrium. The IL-10 anti-inflammatory cytokines, which are produced by central natural killer cells and neutrophils, can interact with the IL-10 receptor on microglia, thereby inhibiting the dysfunction of neurons caused by the over-activation of microglia. It can also inhibit the secretion of TNF by microglia, thereby facilitating the restoration of microglia homeostasis and forming a benign circuit to prevent the further progression of the pathological process.59
Brain Region-Specific Regulation
A variety of inflammatory signals from peripheral inflammation are transmitted to the CNS and subsequently increase neuronal activity in specific regions of the brain. After consolidating across multiple cerebral nuclei, the information is transmitted to the periphery, where it elicits a variety of responses, including anti-inflammatory effects. The hypothalamic-pituitary-adrenal (HPA) axis can be activated by inflammatory signals through a brainstem-hypothalamic pathway of the dorsal vagal complex (DVC)-parabrachial nucleus (PB)-paraventricular nucleus of the hypothalamus(PVN). This pathway leads to the adrenal cortex producing glucocorticoids (GCs) that enter the circulation and produce the immune suppressive effect.44 This effect prevents the persistent elevation of peripheral pro-inflammatory cytokines, which in turn acts on the hypothalamus and pituitary via a feedback effect.60,61 Furthermore, the activation of PVN is accompanied by cognition and illness behaviors, which are the result of the integration of signals from other brain regions.62 Not all pro-inflammatory cytokines cause illness behaviors after signaling to the CNS and the release of IL-17a during inflammation can help improve impairments in social behavior by influencing CNS neuronal activity.63 Furthermore, the information about peripheral inflammation can be retained in the central processing. It has been discovered that certain neurons in the insular cortex are responsible for encoding information related to the beginning of peripheral inflammation, and the reactivation of these neurons can trigger peripheral inflammation through the autonomic nervous system.64
As previously stated, peripheral vagal afferent nerves can transmit peripheral inflammatory signals to the NTS. The NTS, along with the dorsal motor nucleus of the vagus (DMV) and the area postrema (AP), make up the dorsal vagal complex (DVC) (Figure 2). The NTS has direct connections to the rostral ventrolateral medulla (RVLM), where the majority of preganglionic sympathetic neurons are situated.65,66 Neurons that release glutamate in the NTS project to preganglionic sympathetic neurons in the RVLM, leading to the activation of the sympathetic nervous system, which helps counteract inflammation generated by TNFα.67 The AP plays a crucial role in regulating various bodily functions such as the vomit reflex, feeding, metabolism, inflammatory response, stress response, and taste aversions.68 It is capable of receiving vagal input signals and owns the distinct advantage of being located outside the BBB, which allows it to detect and transmit peripheral information. Consequently, when the AP is activated during peripheral inflammation, it may lead to a broad spectrum of sickness behaviors.68,69 The peripheral cholinergic anti-inflammatory pathway can be activated to produce anti-inflammatory effects by stimulating the DMN, which is the source of vagal efferent fibers.70,71 In this way, the DVC functions as an adapter interface, as it simultaneously activates the peripheral anti-inflammatory pathway and upstream brain regions (eg the central amygdala nucleus, striatal bed nucleus of the stria terminalis, paraventricular nucleus of the hypothalamus, and ventral preoptic area) to produce relevant anti-inflammatory effects and immune behaviors.72,73 The excitatory glutamatergic neurons are key members in the adapter action of the DVC.74
 |
Figure 2 The brain’s responses to peripherally derived inflammatory signalling via neural transmission. Abbreviations: Glu, glutamate; DVC, dorsal vagal complex; DRG, dorsal root ganglia; NTS, the nucleus tractus solitarius; DMV, the dorsal motor nucleus of the vagus; AP, area postrema; DRG, dorsal root ganglia; RVLM, rostral ventrolateral medulla. |
In addition, peripheral inflammation induced changes in the rats’ hippocampal glutamate receptors, leading to impaired cognitive function.75 The elevated levels of glutamate in the nucleus ambiguus of mice with sepsis have been shown to exert a neurotoxic effect on cholinergic neurons, ultimately resulting in the inhibition of peripheral cholinergic anti-inflammatory effects.76 This is coupled with the fact that the previously described peripheral inflammatory signals transmitted to the CNS via vagal afferent nerves result in elevated glutamate levels in the CNS. Consequently, these findings underscore the imperative to acknowledge the neurotoxicity associated with the glutamatergic system within the CNS. This phenomenon may also be associated with inflammatory encephalopathies resulting from multiple peripheral inflammatory conditions.77 However, the current study did not explore the changes in extracellular glutamate concentration in different brain regions upon peripheral inflammation. In vivo real-time monitoring of glutamate concentrations is required for a more accurate understanding of their spatiotemporal dynamics.
Central Cholinergic Inhibition and Alterations in Immune Plasticity
Peripheral inflammation not only activates central-associated nerve anti-inflammatory systems but also triggers competing inhibitory effects to achieve a state of balance. The central cholinergic nerves, projecting to the forebrain which contains a high density of M1 muscarinic acetylcholine receptor (M1 mAChR), play an important role in the regulation of peripheral inflammation.78 The selective optogenetic modulation of these forebrain cholinergic neurons has been shown to exert anti-inflammatory effects in the periphery by relying on the structural integrity of the vagus nerve.79,80 This central cholinergic anti-inflammatory modulation would then be relatively weak in diseases associated with cholinergic dysfunction, such as Alzheimer’s disease. Inhibitory effects on the central cholinergic nervous system are induced by peripheral inflammation, which includes the decrease of M1 mAChR and acetylcholine transferase expression and the increase of cholinesterase expression.51,81 However, the selective activation of central cholinergic neurons can alleviate the peripheral inflammation in sepsis mice,79 indicating that its peripheral anti-inflammatory benefits are probably being discounted by these Inhibitory effects.
Cholinergic neurons are primarily located in the basal forebrain and brainstem of the brain. They release acetylcholine through their widely spread axonal fibers, which control neural activity in various nuclei and play a crucial role in functions like locomotion, sleep, emotion, and memory.82,83 In the presence of peripheral inflammation, the population of central cholinergic neurons decreases, as well as the number of cholinergic neurons projecting from the basal forebrain to the hippocampus.84 The hippocampus is a brain region involved in the regulation of cognition and learning. Thus, some researchers have proposed that this reduction of cholinergic neurons in the hippocampus may be the underlying mechanism of neuropsychiatric symptoms caused by peripheral inflammation, and enhancing cholinergic neuron signaling to the hippocampus or increasing hippocampal acetylcholine levels using cholinesterase inhibitors can improve cognitive impairments in mice with sepsis.85–87 Peripheral inflammation has a suppressive impact on the central cholinergic nerves, impairing both cognitive performance and its ability to regulate peripheral anti-inflammatory responses. However, whether these two effects are separate or combined is unclear and requires further attention and investigation.
Moreover, it was discovered that there was an increasing level of chemokine CCL17 produced by the specific group of neurons in the hippocampal CA1 region when mice were injected with LPS.88 The aforementioned implies that the hippocampus is also involved in the entire regulatory chain that contributes to the formation of peripheral inflammation. Interestingly, a study discovered that long-term activation of innate immunity brought on by peripheral inflammation did not alter hippocampal neurological plasticity or cause depressive-like behaviors; rather, hippocampus structure and function could remain intact.89 Perhaps, the innate immunity undergoes remodeling and strengthening throughout chronic activation to adapt to external changes, which is a result of its plasticity,90 this process also serves as a protective mechanism for the CNS. Likewise, the adaptive immune system exhibits plasticity. The CNS becomes desensitized and tolerant to neuroinflammation due to repeated peripheral inflammation, that may persist over time. This tolerance may attenuate neurological hyperreactivity, the activation of microglia, and the expression of inflammatory cytokines in the brain as a result of peripheral inflammation.91,92
Peripheral inflammation leads to an increase in pro-inflammatory cytokine expression in both peripheral and specific regions of the brain.52,53 This is accompanied by immune cell infiltration, degenerative demyelination of neurons, activation of microglia, and destruction of the BBB, all of which contribute to the emergence of central neuroinflammation. In neurodegenerative diseases, such as multiple sclerosis and Parkinson’s disease, if peripheral inflammation persists, it will worsen the primary lesion and further exacerbate cognitive dysfunction and anxiety-like symptoms.93,94 So it is crucial to promptly control peripheral inflammation in these patients.
Overall, it is crucial not to overlook the safeguarding of the CNS in the presence of peripheral inflammation. While previous studies have identified brain regions involved in controlling peripheral inflammation, it is important to note that this does not necessarily indicate a direct connection between these brain regions. There may be additional nuclei or neural pathways responsible for regulating peripheral inflammation or more neural pathways within the highly intricate CNS. The brain’s reaction to peripheral inflammatory signals can be characterized as extensive since it decodes and analyzes the signals sent by the peripheral afferent nerves system, as well as activates other relevant brain regions to allow the body to execute responses other than anti-inflammatory ones, which necessitate a high level of accuracy and promptness.
Peripheral Anti-Inflammatory Pathways
The autonomic nervous system and the neuroendocrine system are the primary mechanisms by which the brain regulates the immune system. The parasympathetic and sympathetic nervous systems are involved in the autonomic response; instead, the HPA axis is mainly involved in the neuroendocrine response (Figure 3).
 |
Figure 3 The neuroimmune circuits in peripheral inflammation. Abbreviations: BBB, blood-brain barrier; PVN, paraventricular nucleus of the hypothalamus; CRH, corticotropin-releasing hormone; ACTH, adrenocorticotropic hormone; GCs, glucocorticoids; DMV, the dorsal motor nucleus of the vagus; NTS, the nucleus tractus solitarius; Ach, acetylcholine; NE, norepinephrine; E, epinephrine; Mφ, macrophage. |
Cholinergic Anti-Inflammatory Pathway
The neuroendocrine HPA axis and autonomic nerves should be relatively independent in their involvement in inflammatory regulation, as the activation of the HPA is not significantly impacted by subphrenic vagotomy.23 In contrast to the vagal anti-inflammatory reflex, which responds swiftly to inflammation, the HPA axis induces a long-lasting anti-inflammatory response throughout the body, though at a slower pace.95 Inflammatory signals are transmitted to the brain through vagal afferent nerves, which immediately activate vagal efferent nerves. These nerves complete neurotransformation at the celiac ganglion and subsequently activate parasympathetic noradrenergic neurons that innervate the spleen to release norepinephrine (NE), which continues to act on the β2 adrenergic receptor on the CD4+ T-cells, leading to the secretion of acetylcholine.96,97 Acetylcholine then acts on the α7nACh receptor on the macrophage, inhibiting the secretion of pro-inflammatory cytokine and exerting an anti-inflammatory effect.98,99 This is a classic peripheral vagal cholinergic anti-inflammatory pathway. Furthermore, activated α7nACh receptors can enhance the contacts between macrophages in the spleen, hence further amplifying the results of the anti-inflammatory effect.100
The spleen is an essential component in this pathway. Spleens that are isolated have the ability to generate acetylcholine on their own, and the cholinergic effect of the spleen is amplified in an adverse environment of inflammation.101 The vagus nerve innervates many tissues or organs, and its anti-inflammatory effects should theoretically be widespread. Thus it’s vital to consider whether other tissues are engaged in this system, even though the spleen plays a major role in the cholinergic anti-inflammatory pathway. Within various organs, vagus efferent nerves have the potential to perform anti-inflammatory effects through the application of certain pathways. For example, it was found that in mice with denervated spleens and deficient T-cells, activated vagal efferent nerves interacted with cholinergic enteric myenteric neurons. These activated cholinergic enteric neurons subsequently inhibited the activity of resident enteric myenteric macrophages expressing α7nACh receptors, thereby attenuating intestinal inflammation and improving postoperative intestinal obstruction.102,103 Conversely, it has been discovered that neither bone marrow nor lymph node cells significantly contribute to the vagus nerve stimulation’s protective impact against renal ischemia.104 Importantly, the study of cholinergic anti-inflammatory pathways that specifically target organs or tissues can enable precise modulation of potential neuroimmune therapeutic targets for treating specific diseases. The protective role of this pathway has already been partially demonstrated in experimental disease models, such as acute kidney injury,104 ARDS,105 pancreatitis,106 and sepsis.107
Sympathetic Nervous System
The immune system is regulated by the sympathetic nervous system(SNS) and peripheral cholinergic anti-inflammatory pathways in a synergistic manner during peripheral inflammation.108 The SNS is composed of two distinct groups of neurons: the preganglionic cholinergic neurons, which secrete acetylcholine, and the postganglionic adrenergic neurons, which primarily secrete NE. These neurons are connected by synaptic contact at the paravertebral nerve nodes. The adrenal medulla is distinguished by the fact that it is the sole recipient of preganglionic sympathetic innervation and secretes NE or epinephrine (E) into the bloodstream to modulate peripheral inflammation.109 In contrast to the parasympathetic nervous system, the majority of immune organs (such as the spleen, lymph nodes, and thymus) receive a significant number of innervations from sympathetic postganglionic neurons.110 There are also adrenergic receptors on innate and adaptive immune cells that possess the ability to bind NE, particularly β2-adrenergic receptors, which enable them to interact directly with the SNS.111,112
The primary mechanism by which the SNS induces an immunosuppressive response is the secretion of the neurotransmitter NE. NE inhibits excessive inflammation in LPS-stimulated mice by dose-dependently reducing the production of pro-inflammatory cytokines in immune cells, simultaneously increasing the production of the anti-inflammatory IL-10 and inhibiting the migration of lymphocytes from lymph nodes to peripheral tissues.113–115 The depletion of peripheral adrenergic nerves by injection of 6-hydroxydopamine hydrochloride (6-OHD) in mice with LPS-induced inflammation reduces the efficacy of inflammation resolution and contributes to a hyperinflammatory state.116 In the same vein, the immunosuppressive impacts of sympathetic activation have been manifested in human clinical trials, and it probably serves as an important driver of immunosuppression in sepsis.117,118 The IL-10 and TNF cytokine responses in mice with intravenous LPS-induced inflammation were unaffected by the combined excision of cervical and lumbar sympathetic nerves, indicating that the endogenous anti-inflammatory effects of sympathetic nerves are primarily derived from the thoracic sympathetic nerves that innervate the abdominal viscera.119
Nevertheless, it is hard to ignore the possibility that the current discoveries about the respective anti-inflammatory effects of the two routes are the consequence of a combination of their actions since NE and β2 adrenergic receptors also contribute to the cholinergic anti-inflammatory pathway. The SNS may be more advantageous for immune regulation considering the immune system’s innervation and receptor distribution. However, its activation as the basis for the body’s “fight-or-flight” response is nonspecific and spontaneous (especially in disease). And pain, stress, or physiological changes in the body can increase its tone.120,121 There is a significant risk of sympatho-vagal imbalance with continuous sympathetic activation and vagal inhibition, resulting in irreversible immunosuppression, if the nerve tone of the SNS is continuously increased to achieve immunomodulation. This autonomic dysfunction will exacerbate the severity of the disease and result in an unfavorable prognosis.117,122,123 Controlling the hypertonic SNS to achieve sympatho-vagal rebalance can thus have a therapeutic effect. Dextromethorphan can prevent sympathetic hyperactivation and cause a relative rise in peripheral cholinergic vagal anti-inflammatory nerve tone in a mouse model of experimental severe acute pancreatitis, which in turn, lowers systemic inflammation and pancreatic damage.124 In clinical trials, thoracic epidural analgesia decreases mortality, enhances patient prognosis, and controls sympatho-vagal imbalance caused by acute pancreatitis.125,126 As a systemic inflammatory treatment, it is not a bad notion to stabilize autonomic homeostasis by controlling over-activated sympathetic nerves, in addition to treating peripheral inflammation by stimulating the vagus nerve (Table 1).
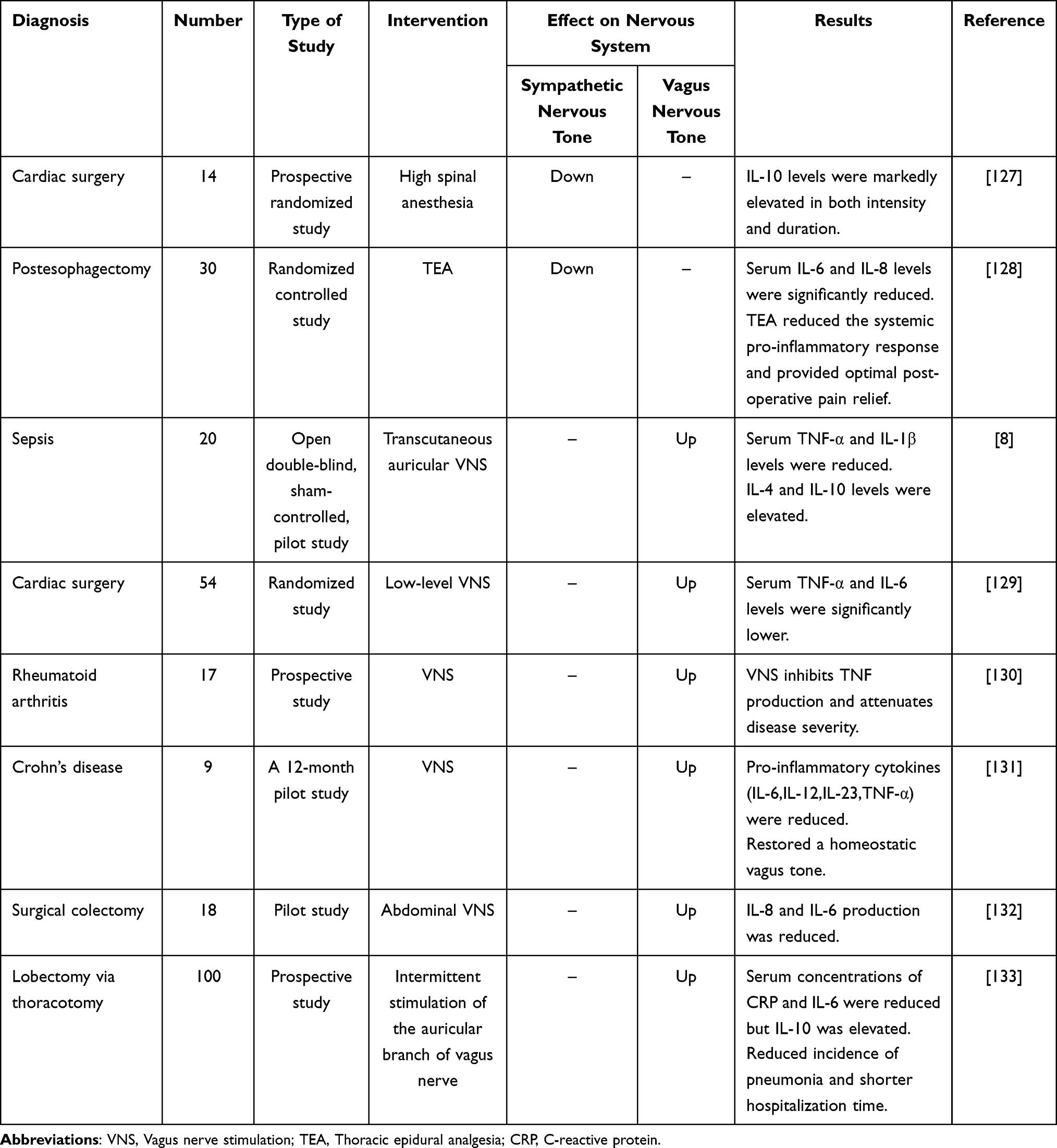 |
Table 1 Inflammation Treatment by Intervening Nerve System in Humans |
Vagal-Adrenal Anti-Inflammatory Pathway
In addition to the two anti-inflammatory pathways previously mentioned, a vagal-adrenal anti-inflammatory pathway has been recently identified which induces a protective neurological response in rodents by electrically stimulating vagal efferent fibers in the absence of afferent vagal signals.104,134 Specific peripheral somatosensory receptors may be present in this pathway. In contrast to the electrical stimulation of spinal sympathetic reflexes which are not dependent on the presence of the PROKR2ADV neurons, Shenbin Liu and Zhifu Wang discovered that selectively stimulating sensory neurons expressing Prokr2 protein, which is distributed somatosensory-specifically (innervating deep fascial tissues such as periosteum, articular ligaments, and myofascia in the limbs, but not the epidermal tissues of the skin and major fascial tissues such as the peritoneum), would activate a vagal-adrenal anti-inflammatory pathway to inhibit inflammatory storms caused by the LPS treatment.134,135
In response to systemic inflammation, each of these anti-inflammatory pathways can be activated to carry out immunomodulatory functions. While they may appear to be independent, the immune system can be influenced by various interactions between them. In general, the activation of these pathways can be identified as a synergistic effect of immunosuppression. However, each pathway will achieve an overall balance through its feedback or regulation. If the balance among anti-inflammatory pathways is abnormal, it may result in a poor prognosis owing to irremediable immunosuppression in the later stages of the disease. Therefore, neurological interventions can be used in the early stages to control the dysregulated neurological balance.
Conclusion
This review offers a macroscopic perspective on the neuroimmune communication that occurs during peripheral inflammation. Peripheral inflammation initiates this reflex by transmitting sensory information to the spinal cord and brain via multiple pathways. This process subsequently stimulates the peripheral autonomic nervous system or the release of hormones, resulting in the modulation of various immune system functions. The altered peripheral inflammatory signals continue to be transmitted upwards, forming a closed loop. Inflammation is accompanied by a series of disagreeable sensations and disease behaviors due to the presence of numerous branches of central regulation in a closed-loop regulation process. Even though a growing body of research is gradually extending our understanding of neuroimmune and increasing the number of neuroimmune junctions that can be investigated as potential therapeutic targets (eg, interventions for increased BBB permeability, microglia activation, neural pathway imbalances), more animal and clinical studies are needed to develop definitive neuroimmune therapies for peripheral inflammation that are less harmful to the patient, early, and short-cycle.
Funding
This study was supported by grants from the National Natural Science Foundation (No. 8236031060).
Disclosure
No author has financial or other contractual agreements that might cause conflicts of interest.
References
1. Tracey KJ. The inflammatory reflex. Nature. 2002;420(6917):853–859. doi:10.1038/nature01321
2. Sendler M, Van Den Brandt C, Glaubitz J, et al. NLRP3 Inflammasome Regulates Development of Systemic Inflammatory Response and Compensatory Anti-Inflammatory Response Syndromes in Mice With Acute Pancreatitis. Gastroenterology. 2020;158(1):253–269.e14. doi:10.1053/j.gastro.2019.09.040
3. Adib-Conquy M, Cavaillon JM. Compensatory anti-inflammatory response syndrome. Thromb Haemost. 2009;101(1):36–47. doi:10.1160/TH08-07-0421
4. Borovikova LV, Ivanova S, Zhang M, et al. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature. 2000;405(6785):458–462. doi:10.1038/35013070
5. Huang S, Ziegler CGK, Austin J, et al. Lymph nodes are innervated by a unique population of sensory neurons with immunomodulatory potential. Cell. 2021;184(2):441–459.e25. doi:10.1016/j.cell.2020.11.028
6. Ulloa L. Electroacupuncture activates neurons to switch off inflammation. Nature. 2021;598(7882):573–574. doi:10.1038/d41586-021-02714-0
7. Donegà M, Fjordbakk CT, Kirk J, et al. Human-relevant near-organ neuromodulation of the immune system via the splenic nerve. Proc Natl Acad Sci. 2021;118(20):e2025428118. doi:10.1073/pnas.2025428118
8. Wu Z, Zhang X, Cai T, et al. Transcutaneous auricular vagus nerve stimulation reduces cytokine production in sepsis: an open double-blind, sham-controlled, pilot study. Brain Stimul. 2023;16(2):507–514. doi:10.1016/j.brs.2023.02.008
9. Okusa MD, Rosin DL, Tracey KJ. Targeting neural reflex circuits in immunity to treat kidney disease. Nat Rev Nephrol. 2017;13(11):669–680. doi:10.1038/nrneph.2017.132
10. Takeuchi O, Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140(6):805–820. doi:10.1016/j.cell.2010.01.022
11. Steinberg BE, Silverman HA, Robbiati S, et al. Cytokine-specific Neurograms in the Sensory Vagus Nerve. Bioelectron Med. 2016;3(1):7–17. doi:10.15424/bioelectronmed.2016.00007
12. Zanos TP, Silverman HA, Levy T, et al. Identification of cytokine-specific sensory neural signals by decoding murine vagus nerve activity. Proc Natl Acad Sci. 2018;115(21):E4843–E4852. doi:10.1073/pnas.1719083115
13. Jin H, Li M, Jeong E, Castro-Martinez F, Zuker CS. A body-brain circuit that regulates body inflammatory responses. Nature. 2024;630(8017):695–703. doi:10.1038/s41586-024-07469-y
14. Mascarucci P, Perego C, Terrazzino S, De Simoni MG. Glutamate release in the nucleus tractus solitarius induced by peripheral lipopolysaccharide and interleukin-1 beta. Neuroscience. 1998;86(4):1285–1290. doi:10.1016/S0306-4522(98)00105-5
15. Mazzone SB, Undem BJ. Vagal Afferent Innervation of the Airways in Health and Disease. Physiol Rev. 2016;96(3):975–1024. doi:10.1152/physrev.00039.2015
16. Alim MA, Grujic M, Ackerman PW, et al. Glutamate triggers the expression of functional ionotropic and metabotropic glutamate receptors in mast cells. Cell Mol Immunol. 2021;18(10):2383–2392. doi:10.1038/s41423-020-0421-z
17. Yang J, Dong HQ, Liu YH, et al. Laparotomy-Induced Peripheral Inflammation Activates NR2B Receptors on the Brain Mast Cells and Results in Neuroinflammation in a Vagus Nerve-Dependent Manner. Front Cell Neurosci. 2022;16:771156. doi:10.3389/fncel.2022.771156
18. Verma M, Lizama BN, Chu CT. Excitotoxicity, calcium and mitochondria: a triad in synaptic neurodegeneration. Transl Neurodegener. 2022;11(1):3. doi:10.1186/s40035-021-00278-7
19. Koivisto AP, Belvisi MG, Gaudet R, Szallasi A. Advances in TRP channel drug discovery: from target validation to clinical studies. Nat Rev Drug Discovery. 2022;21(1):41–59. doi:10.1038/s41573-021-00268-4
20. Silverman HA, Tynan A, Hepler TD, et al. Transient Receptor Potential Ankyrin-1-expressing vagus nerve fibers mediate IL-1β induced hypothermia and reflex anti-inflammatory responses. Mol Med. 2023;29(1):4. doi:10.1186/s10020-022-00590-6
21. Joffre J, Wong E, Lawton S, et al. N-Oleoyl dopamine induces IL-10 via central nervous system TRPV1 and improves endotoxemia and sepsis outcomes. J Neuroinflammation. 2022;19(1):118. doi:10.1186/s12974-022-02485-z
22. Yj L, Rl L, Ruan T, Khosravi M, Lee LY. A synergistic effect of simultaneous TRPA1 and TRPV1 activations on vagal pulmonary C-fiber afferents. J Appl Physiol. 2015;118(3):273–281. doi:10.1152/japplphysiol.00805.2014
23. Kobrzycka A, Napora P, Pearson BL, Pierzchała-Koziec K, Szewczyk R, Wieczorek M. Peripheral and central compensatory mechanisms for impaired vagus nerve function during peripheral immune activation. J Neuroinflammation. 2019;16(1):150. doi:10.1186/s12974-019-1544-y
24. Liebner S, Dijkhuizen RM, Reiss Y, Plate KH, Agalliu D, Constantin G. Functional morphology of the blood-brain barrier in health and disease. Acta Neuropathol. 2018;135(3):311–336. doi:10.1007/s00401-018-1815-1
25. Yazdani S, Jaldin-Fincati JR, Pereira RVS, Klip A. Endothelial cell barriers: transport of molecules between blood and tissues. Traffic. 2019;20(6):390–403. doi:10.1111/tra.12645
26. Prinz M, Priller J. The role of peripheral immune cells in the CNS in steady state and disease. Nat Neurosci. 2017;20(2):136–144. doi:10.1038/nn.4475
27. Beltran-Velasco AI, Clemente-Suárez VJ. Impact of peripheral inflammation on blood-brain barrier dysfunction and its role in neurodegenerative diseases. Int J Mol Sci. 2025;26(6):2440. doi:10.3390/ijms26062440
28. Huang X, Hussain B, Chang J. Peripheral inflammation and blood-brain barrier disruption: effects and mechanisms. CNS Neurosci Ther. 2021;27(1):36–47. doi:10.1111/cns.13569
29. Banks WA, Hansen KM, Erickson MA, Crews FT. High-mobility group box 1 (HMGB1) crosses the BBB bidirectionally. Brain Behav Immun. 2023;111:386–394. doi:10.1016/j.bbi.2023.04.018
30. Da Rocha GHO, Loiola RA, M DP-S, et al. Pioglitazone Attenuates the Effects of Peripheral Inflammation in a Human In Vitro Blood-Brain Barrier Model. Int J Mol Sci. 2022;23(21):12781. doi:10.3390/ijms232112781
31. Shaker T, Chattopadhyaya B, Amilhon B, Cristo GD, Weil AG. Transduction of inflammation from peripheral immune cells to the hippocampus induces neuronal hyperexcitability mediated by Caspase-1 activation. Neurobiol Dis. 2021;160:105535. doi:10.1016/j.nbd.2021.105535
32. Spiteri AG, Wishart CL, Pamphlett R, Locatelli G, King NJC. Microglia and monocytes in inflammatory CNS disease: integrating phenotype and function. Acta Neuropathol. 2022;143(2):179–224.
33. Xie X, Luo X, Liu N, et al. Monocytes, microglia, and CD200-CD200R1 signaling are essential in the transmission of inflammation from the periphery to the central nervous system. J Neurochem. 2017;141(2):222–235. doi:10.1111/jnc.13972
34. Ruiz VY, Calderon TM, Leon-Rivera R, Chilunda V, Zhang J, Berman JW. Single-cell analysis of CD14+CD16+ monocytes identifies a subpopulation with an enhanced migratory and inflammatory phenotype. Front Immunol. 2025;16:1475480. doi:10.3389/fimmu.2025.1475480
35. Li C, Niu C, Chen L, et al. Classical monocytes shuttling for precise delivery of nanotherapeutics to glioblastoma. Adv Healthcare Mater. 2024;13(29):e2400925. doi:10.1002/adhm.202400925
36. Cheng R, Wang S. Cell-mediated nanoparticle delivery systems: towards precision nanomedicine. Drug Deliv Transl Res. 2024;14(11):3032–3054. doi:10.1007/s13346-024-01591-0
37. Filippi M, Bar-Or A, Piehl F, et al. Multiple sclerosis. Nat Rev Dis Primers. 2018;4(1):43. doi:10.1038/s41572-018-0041-4
38. Korn T, Kallies A. T cell responses in the central nervous system. Nat Rev Immunol. 2017;17(3):179–194. doi:10.1038/nri.2016.144
39. Duan L, Zhang XD, Miao WY, et al. PDGFRβ Cells Rapidly Relay Inflammatory Signal from the Circulatory System to Neurons via Chemokine CCL2. Neuron. 2018;100(1):183–200.e8. doi:10.1016/j.neuron.2018.08.030
40. Fricke F, Gebert J, Kopitz J, Plaschke K. Proinflammatory extracellular vesicle-mediated signaling contributes to the induction of neuroinflammation in animal models of endotoxemia and peripheral surgical stress. Cell Mol Neurobiol. 2021;41(6):1325–1336. doi:10.1007/s10571-020-00905-3
41. Balusu S, Van Wonterghem E, De Rycke R, et al. Identification of a novel mechanism of blood-brain communication during peripheral inflammation via choroid plexus-derived extracellular vesicles. EMBO Mol Med. 2016;8(10):1162–1183. doi:10.15252/emmm.201606271
42. Lazarevic I, Soldati S, Mapunda JA, et al. The choroid plexus acts as an immune cell reservoir and brain entry site in experimental autoimmune encephalomyelitis. Fluids Barriers CNS. 2023;20(1):39. doi:10.1186/s12987-023-00441-4
43. Dantzer R. Cytokine-induced sickness behavior: where do we stand? Brain Behav Immun. 2001;15(1):7–24. doi:10.1006/brbi.2000.0613
44. Jagot F, Gaston-Breton R, Choi AJ, et al. The parabrachial nucleus elicits a vigorous corticosterone feedback response to the pro-inflammatory cytokine IL-1β. Neuron. 2023;111(15):2367–2382.e6. doi:10.1016/j.neuron.2023.05.009
45. Borst K, Dumas AA, Prinz M. Microglia: immune and non-immune functions. Immunity. 2021;54(10):2194–2208. doi:10.1016/j.immuni.2021.09.014
46. Castro LVG, Gonçalves-de-Albuquerque CF, Silva AR. Polarization of Microglia and Its Therapeutic Potential in Sepsis. Int J Mol Sci. 2022;23(9):4925. doi:10.3390/ijms23094925
47. Riester K, Brawek B, Savitska D, et al. In vivo characterization of functional states of cortical microglia during peripheral inflammation. Brain Behav Immun. 2020;87:243–255. doi:10.1016/j.bbi.2019.12.007
48. Nayak D, Roth TL, McGavern DB. Microglia development and function. Annu Rev Immunol. 2014;32(1):367–402. doi:10.1146/annurev-immunol-032713-120240
49. Barbosa-Silva MC, Rm C, Del Castilo I, et al. Mice lacking 5-lipoxygenase display motor deficits associated with cortical and hippocampal synapse abnormalities. Brain Behav Immun. 2022;100:183–193. doi:10.1016/j.bbi.2021.12.004
50. Di Filippo M, Chiasserini D, Gardoni F, et al. Effects of central and peripheral inflammation on hippocampal synaptic plasticity. Neurobiol Dis. 2013;52:229–236. doi:10.1016/j.nbd.2012.12.009
51. Zaghloul N, Addorisio ME, Silverman HA, et al. Forebrain Cholinergic Dysfunction and Systemic and Brain Inflammation in Murine Sepsis Survivors. Front Immunol. 2017;8:1673. doi:10.3389/fimmu.2017.01673
52. Silverman HA, Dancho M, Regnier-Golanov A, et al. Brain region-specific alterations in the gene expression of cytokines, immune cell markers and cholinergic system components during peripheral endotoxin-induced inflammation. Mol Med. 2014;20(1):601–611. doi:10.2119/molmed.2014.00147
53. Qin L, Wu X, Block ML, et al. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia. 2007;55(5):453–462. doi:10.1002/glia.20467
54. García-Domínguez I, Veselá K, García-Revilla J, et al. Peripheral Inflammation Enhances Microglia Response and Nigral Dopaminergic Cell Death in an in vivo MPTP Model of Parkinson’s Disease. Front Cell Neurosci. 2018;12:398. doi:10.3389/fncel.2018.00398
55. Ho YH, Lin YT, Wu CWJ, Chao YM, Chang AYW, Chan JYH. Peripheral inflammation increases seizure susceptibility via the induction of neuroinflammation and oxidative stress in the hippocampus. J Biomed Sci. 2015;22(1):46. doi:10.1186/s12929-015-0157-8
56. Lin J, Xu Y, Guo P, et al. CCL5/CCR5-mediated peripheral inflammation exacerbates blood‒brain barrier disruption after intracerebral hemorrhage in mice. J Transl Med. 2023;21(1):196. doi:10.1186/s12967-023-04044-3
57. Da Costa LHA, Santos-Junior NN, Catalão CHR, Rocha MJA. Microglial Activation Modulates Neuroendocrine Secretion During Experimental Sepsis. Mol Neurobiol. 2021;58(5):2133–2144. doi:10.1007/s12035-020-02241-5
58. Riazi K, Galic MA, Kentner AC, Reid AY, Sharkey KA, Pittman QJ. Microglia-dependent alteration of glutamatergic synaptic transmission and plasticity in the hippocampus during peripheral inflammation. J Neurosci. 2015;35(12):4942–4952. doi:10.1523/JNEUROSCI.4485-14.2015
59. Shemer A, Scheyltjens I, Frumer GR, et al. Interleukin-10 Prevents Pathological Microglia Hyperactivation following Peripheral Endotoxin Challenge. Immunity. 2020;53(5):1033–1049.e7. doi:10.1016/j.immuni.2020.09.018
60. Cain DW, Cidlowski JA. Immune regulation by glucocorticoids. Nat Rev Immunol. 2017;17(4):233–247. doi:10.1038/nri.2017.1
61. Keller-Wood M. Hypothalamic-Pituitary--Adrenal Axis-Feedback Control. Compr Physiol. 2015;5(3):1161–1182. doi:10.1002/j.2040-4603.2015.tb00644.x
62. Granton E, Brown L, Defaye M, et al. Biofilm exopolysaccharides alter sensory-neuron-mediated sickness during lung infection. Cell. 2024;187(8):1874–1888.e14. doi:10.1016/j.cell.2024.03.001
63. Reed MD, Yim YS, Wimmer RD, et al. IL-17a promotes sociability in mouse models of neurodevelopmental disorders. Nature. 2020;577(7789):249–253. doi:10.1038/s41586-019-1843-6
64. Koren T, Yifa R, Amer M, et al. Insular cortex neurons encode and retrieve specific immune responses. Cell. 2021;184(24):5902–5915.e17. doi:10.1016/j.cell.2021.10.013
65. Farmer DGS, Pracejus N, Dempsey B, et al. On the presence and functional significance of sympathetic premotor neurons with collateralized spinal axons in the rat. J Physiol. 2019;597(13):3407–3423. doi:10.1113/JP277661
66. Kline DD, King TL, Austgen JR, Heesch CM, Hasser EM. Sensory afferent and hypoxia-mediated activation of nucleus tractus solitarius neurons that project to the rostral ventrolateral medulla. Neuroscience. 2010;167(2):510–527. doi:10.1016/j.neuroscience.2010.02.012
67. Katayama PL, Leirão IP, Kanashiro A, et al. The carotid body detects circulating tumor necrosis factor-alpha to activate a sympathetic anti-inflammatory reflex. Brain Behav Immun. 2022;102:370–386. doi:10.1016/j.bbi.2022.03.014
68. Cheng W, Gordian D, Ludwig MQ, Pers TH, Seeley RJ, Myers MG. Hindbrain circuits in the control of eating behaviour and energy balance. Nat Metab. 2022;4(7):826–835. doi:10.1038/s42255-022-00606-9
69. Zhang C, Kaye JA, Cai Z, Wang Y, Prescott SL, Liberles SD. Area Postrema Cell Types that Mediate Nausea-Associated Behaviors. Neuron. 2021;109(3):461–472.e5. doi:10.1016/j.neuron.2020.11.010
70. Kressel AM, Tsaava T, Levine YA, et al. Identification of a brainstem locus that inhibits tumor necrosis factor. Proc Natl Acad Sci. 2020;117(47):29803–29810. doi:10.1073/pnas.2008213117
71. Thompson DA, Tsaava T, Rishi A, et al. Optogenetic stimulation of the brainstem dorsal motor nucleus ameliorates acute pancreatitis. Front Immunol. 2023;14:1166212. doi:10.3389/fimmu.2023.1166212
72. Marvel FA, Chen CC, Badr N, Gaykema RPA, Goehler LE. Reversible inactivation of the dorsal vagal complex blocks lipopolysaccharide-induced social withdrawal and c-Fos expression in central autonomic nuclei. Brain Behav Immun. 2004;18(2):123–134. doi:10.1016/j.bbi.2003.09.004
73. Wean JB, Smith BN. Fibroblast Growth Factor 19 Increases the Excitability of Pre-Motor Glutamatergic Dorsal Vagal Complex Neurons From Hyperglycemic Mice. Front Endocrinol. 2021;12:765359. doi:10.3389/fendo.2021.765359
74. Clyburn C, Browning KN. Glutamatergic plasticity within neurocircuits of the dorsal vagal complex and the regulation of gastric functions. Am J Physiol Gastr Liver Physiol. 2021;320(5):G880–G887.
75. Balzano T, Dadsetan S, Forteza J, et al. Chronic hyperammonemia induces peripheral inflammation that leads to cognitive impairment in rats: reversed by anti-TNF-α treatment. J Hepatol. 2020;73(3):582–592. doi:10.1016/j.jhep.2019.01.008
76. Jiang J, Ai S, Yuan C, et al. Dysfunction of cholinergic neuron in nucleus ambiguous aggravates sepsis-induced lung injury via a GluA1-dependment mechanism. Brain Behav Immun. 2025;128:689–702. doi:10.1016/j.bbi.2025.05.006
77. Mei B, Xu X, Weng J, et al. Activating astrocytic α2A adrenoceptors in hippocampus reduces glutamate toxicity to attenuate sepsis-associated encephalopathy in mice. Brain Behav Immun. 2024;117:376–398. doi:10.1016/j.bbi.2024.02.005
78. Lehner KR, Silverman HA, Addorisio ME, et al. Forebrain Cholinergic Signaling Regulates Innate Immune Responses and Inflammation. Front Immunol. 2019;10:585. doi:10.3389/fimmu.2019.00585
79. Zhai Q, Lai D, Cui P, et al. Selective activation of basal forebrain cholinergic neurons attenuates polymicrobial Sepsis–Induced inflammation via the cholinergic anti-inflammatory pathway. Crit Care Med. 2017;45(10):e1075–e1082. doi:10.1097/CCM.0000000000002646
80. Rosas-Ballina M, Valdés-Ferrer SI, Dancho ME, et al. Xanomeline suppresses excessive pro-inflammatory cytokine responses through neural signal-mediated pathways and improves survival in lethal inflammation. Brain Behav Immun. 2015;44:19–27. doi:10.1016/j.bbi.2014.07.010
81. Abraham MN, Nedeljkovic-Kurepa A, Fernandes TD, et al. M1 cholinergic signaling in the brain modulates cytokine levels and splenic cell sub-phenotypes following cecal ligation and puncture. Mol Med. 2024;30(1):22. doi:10.1186/s10020-024-00787-x
82. Li X, Yu B, Sun Q, et al. Generation of a whole-brain atlas for the cholinergic system and mesoscopic projectome analysis of basal forebrain cholinergic neurons. Proc Natl Acad Sci. 2018;115(2):415–420. doi:10.1073/pnas.1703601115
83. Ananth MR, Rajebhosale P, Kim R, Talmage DA, Role LW. Basal forebrain cholinergic signalling: development, connectivity and roles in cognition. Nat Rev Neurosci. 2023;24(4):233–251. doi:10.1038/s41583-023-00677-x
84. He G, Li Y, Deng H, Zuo H. Advances in the study of cholinergic circuits in the central nervous system. Ann Clin Transl Neurol. 2023;10(12):2179–2191. doi:10.1002/acn3.51920
85. Liu Y, Zhang Y, Zheng X, et al. Galantamine improves cognition, hippocampal inflammation, and synaptic plasticity impairments induced by lipopolysaccharide in mice. J Neuroinflammation. 2018;15(1):112. doi:10.1186/s12974-018-1141-5
86. Y L, Z J, M H, et al. Selective activation of cholinergic neurotransmission from the medial septal nucleus to hippocampal pyramidal neurones improves sepsis-induced cognitive deficits in mice. Br J Anaesth. 2023;130(5):1.
87. Suarez AN, Hsu TM, Liu CM, et al. Gut vagal sensory signaling regulates hippocampus function through multi-order pathways. Nat Commun. 2018;9(1):2181. doi:10.1038/s41467-018-04639-1
88. Fülle L, Offermann N, Hansen JN, et al. CCL17 exerts a neuroimmune modulatory function and is expressed in hippocampal neurons. Glia. 2018;66(10):2246–2261. doi:10.1002/glia.23507
89. Süß P, Kalinichenko L, Baum W, et al. Hippocampal structure and function are maintained despite severe innate peripheral inflammation. Brain Behav Immun. 2015;49:156–170. doi:10.1016/j.bbi.2015.05.011
90. Florentino D, Della Giustina A, De Souza Goldim MP, et al. Early life neuroimmune challenge protects the brain after sepsis in adult rats. Neurochem Int. 2020;135:104712. doi:10.1016/j.neuint.2020.104712
91. Peng J, Sy CB, Ponessa JJ, et al. Monocytes maintain central nervous system homeostasis following helminth-induced inflammation. Proc Natl Acad Sci. 2022;119(37):e2201645119. doi:10.1073/pnas.2201645119
92. Liu X, Nemeth DP, Tarr AJ, et al. Euflammation attenuates peripheral inflammation-induced neuroinflammation and mitigates immune-to-brain signaling. Brain Behav Immun. 2016;54:140–148. doi:10.1016/j.bbi.2016.01.018
93. Silva BA, Farías MI, Miglietta EA, et al. Understanding the role of the blood brain barrier and peripheral inflammation on behavior and pathology on ongoing confined cortical lesions. Mult Scler Relat Disord. 2022;57:103346. doi:10.1016/j.msard.2021.103346
94. Berriat F, Lobsiger CS, Boillée S. The contribution of the peripheral immune system to neurodegeneration. Nat Neurosci. 2023;26(6):942–954. doi:10.1038/s41593-023-01323-6
95. Chavan SS, Tracey KJ. Essential Neuroscience in Immunology. J Immunol. 2017;198(9):3389–3397. doi:10.4049/jimmunol.1601613
96. Rosas-Ballina M, Ochani M, Parrish WR, et al. Splenic nerve is required for cholinergic antiinflammatory pathway control of TNF in endotoxemia. Proc Natl Acad Sci. 2008;105(31):11008–11013. doi:10.1073/pnas.0803237105
97. Rosas-Ballina M, Olofsson PS, Ochani M, et al. Acetylcholine-synthesizing T cells relay neural signals in a vagus nerve circuit. Science. 2011;334(6052):98–101. doi:10.1126/science.1209985
98. Browning KN, Verheijden S, Boeckxstaens GE. The Vagus Nerve in Appetite Regulation, Mood, and Intestinal Inflammation. Gastroenterology. 2017;152(4):730–744. doi:10.1053/j.gastro.2016.10.046
99. Wang H, Yu M, Ochani M, et al. Nicotinic acetylcholine receptor alpha7 subunit is an essential regulator of inflammation. Nature. 2003;421(6921):384–388. doi:10.1038/nature01339
100. Nakamura Y, Matsumoto H, Wu CH, et al. Alpha 7 nicotinic acetylcholine receptors signaling boosts cell-cell interactions in macrophages effecting anti-inflammatory and organ protection. Commun Biol. 2023;6(1):666. doi:10.1038/s42003-023-05051-2
101. Hoover DB, Poston MD, Brown S, et al. Cholinergic leukocytes in sepsis and at the neuroimmune junction in the spleen. Int Immunopharmacol. 2020;81:106359. doi:10.1016/j.intimp.2020.106359
102. Stakenborg N, Labeeuw E, Gomez-Pinilla PJ, et al. Preoperative administration of the 5-HT4 receptor agonist prucalopride reduces intestinal inflammation and shortens postoperative ileus via cholinergic enteric neurons. Gut. 2019;68(8):1406–1416. doi:10.1136/gutjnl-2018-317263
103. Matteoli G, Gomez-Pinilla PJ, Nemethova A, et al. A distinct vagal anti-inflammatory pathway modulates intestinal muscularis resident macrophages independent of the spleen. Gut. 2014;63(6):938–948. doi:10.1136/gutjnl-2013-304676
104. Tanaka S, Abe C, Abbott SBG, et al. Vagus nerve stimulation activates two distinct neuroimmune circuits converging in the spleen to protect mice from kidney injury. Proc Natl Acad Sci. 2021;118(12):e2021758118. doi:10.1073/pnas.2021758118
105. Zhang X, Wei X, Deng Y, et al. Mesenchymal stromal cells alleviate acute respiratory distress syndrome through the cholinergic anti-inflammatory pathway. Signal Transduct Target Ther. 2022;7(1):307. doi:10.1038/s41392-022-01124-6
106. Van Westerloo DJ, Giebelen IA, Florquin S, et al. The vagus nerve and nicotinic receptors modulate experimental pancreatitis severity in mice. Gastroenterology. 2006;130(6):1822–1830. doi:10.1053/j.gastro.2006.02.022
107. Huston JM, Ochani M, Rosas-Ballina M, et al. Splenectomy inactivates the cholinergic antiinflammatory pathway during lethal endotoxemia and polymicrobial sepsis. J Exp Med. 2006;203(7):1623–1628. doi:10.1084/jem.20052362
108. Bellocchi C, Carandina A, Montinaro B, et al. The Interplay between Autonomic Nervous System and Inflammation across Systemic Autoimmune Diseases. Int J Mol Sci. 2022;23(5):2449. doi:10.3390/ijms23052449
109. Kim HW, Uh DK, Yoon SY, et al. Low-frequency electroacupuncture suppresses carrageenan-induced paw inflammation in mice via sympathetic post-ganglionic neurons, while high-frequency EA suppression is mediated by the sympathoadrenal medullary axis. Brain Res Bull. 2008;75(5):698–705. doi:10.1016/j.brainresbull.2007.11.015
110. Nance DM, Sanders VM. Autonomic innervation and regulation of the immune system (1987–2007). Brain Behav Immun. 2007;21(6):736–745. doi:10.1016/j.bbi.2007.03.008
111. Sanders VM, Straub RH. Norepinephrine, the beta-adrenergic receptor, and immunity. Brain Behav Immun. 2002;16(4):290–332. doi:10.1006/brbi.2001.0639
112. Sharma D, Farrar JD. Adrenergic regulation of immune cell function and inflammation. Semin Immunopathol. 2020;42(6):709–717. doi:10.1007/s00281-020-00829-6
113. Nakai A, Hayano Y, Furuta F, Noda M, Suzuki K. Control of lymphocyte egress from lymph nodes through β2-adrenergic receptors. J Exp Med. 2014;211(13):2583–2598. doi:10.1084/jem.20141132
114. Stolk RF, Van Der Pasch E, Naumann F, et al. Norepinephrine Dysregulates the Immune Response and Compromises Host Defense during Sepsis. Am J Respir Crit Care Med. 2020;202(6):830–842. doi:10.1164/rccm.202002-0339OC
115. Thoppil J, Mehta P, Bartels B, Sharma D, Farrar JD. Impact of norepinephrine on immunity and oxidative metabolism in sepsis. Front Immunol. 2023;14:1271098. doi:10.3389/fimmu.2023.1271098
116. Körner A, Schlegel M, Kaussen T, et al. Sympathetic nervous system controls resolution of inflammation via regulation of repulsive guidance molecule A. Nat Commun. 2019;10(1):633. doi:10.1038/s41467-019-08328-5
117. Stolk RF, Kox M, Pickkers P. Noradrenaline drives immunosuppression in sepsis: clinical consequences. Intensive Care Med. 2020;46(6):1246–1248. doi:10.1007/s00134-020-06025-2
118. Kox M, Van Eijk LT, Zwaag J, et al. Voluntary activation of the sympathetic nervous system and attenuation of the innate immune response in humans. Proc Natl Acad Sci. 2014;111(20):7379–7384. doi:10.1073/pnas.1322174111
119. McKinley MJ, Martelli D, Trevizan‐Baú P, McAllen RM. Divergent splanchnic sympathetic efferent nerve pathways regulate interleukin-10 and tumour necrosis factor-α responses to endotoxaemia. J Physiol. 2022;600(20):4521–4536. doi:10.1113/JP283217
120. Brognara F, Castania JA, Kanashiro A, Dias DPM, Salgado HC. Physiological Sympathetic Activation Reduces Systemic Inflammation: role of Baroreflex and Chemoreflex. Front Immunol. 2021;12:637845. doi:10.3389/fimmu.2021.637845
121. Meltzer JC, MacNeil BJ, Sanders V, et al. Stress-induced suppression of in vivo splenic cytokine production in the rat by neural and hormonal mechanisms. Brain Behav Immun. 2004;18(3):262–273. doi:10.1016/j.bbi.2003.09.003
122. Zhang Y, Popović ZB, Bibevski S, et al. Chronic vagus nerve stimulation improves autonomic control and attenuates systemic inflammation and heart failure progression in a canine high-rate pacing model. Circ Heart Fail. 2009;2(6):692–699. doi:10.1161/CIRCHEARTFAILURE.109.873968
123. Koopman FA, Stoof SP, Straub RH, Van Maanen MA, Vervoordeldonk MJ, Tak PP. Restoring the balance of the autonomic nervous system as an innovative approach to the treatment of rheumatoid arthritis. Mol Med. 2011;17(9–10):937–948. doi:10.2119/molmed.2011.00065
124. Huang DY, Li Q, Shi CY, Hou CQ, Miao Y, Shen HB. Dexmedetomidine attenuates inflammation and pancreatic injury in a rat model of experimental severe acute pancreatitis via cholinergic anti-inflammatory pathway. Chin Med J. 2020;133(9):1073–1079. doi:10.1097/CM9.0000000000000766
125. Jabaudon M, Belhadj-Tahar N, Rimmelé T, et al. Thoracic epidural analgesia and mortality in acute pancreatitis: a multicenter propensity analysis. Crit Care Med. 2018;46(3):e198–e205. doi:10.1097/CCM.0000000000002874
126. Wang Q, Fu B, Su D, Fu X. Impact of early thoracic epidural analgesia in patients with severe acute pancreatitis. Eur J Clin Invest. 2022;52(6):e13740. doi:10.1111/eci.13740
127. Lee TWR, Kowalski S, Falk K, Maguire D, Freed DH, HayGlass KT. High Spinal Anesthesia Enhances Anti-Inflammatory Responses in Patients Undergoing Coronary Artery Bypass Graft Surgery and Aortic Valve Replacement: randomized Pilot Study. PLoS One. 2016;11(3):e0149942. doi:10.1371/journal.pone.0149942
128. Fares KM, Mohamed SA, Hamza HM, Sayed DM, Hetta DF. Effect of thoracic epidural analgesia on pro-inflammatory cytokines in patients subjected to protective lung ventilation during Ivor Lewis esophagectomy. Pain Physician. 2014;17(4):305–315.
129. Stavrakis S, Humphrey MB, Scherlag B, et al. Low-Level Vagus Nerve Stimulation Suppresses Post-Operative Atrial Fibrillation and Inflammation: a Randomized Study. JACC Clin Electrophysiol. 2017;3(9):929–938. doi:10.1016/j.jacep.2017.02.019
130. Koopman FA, Chavan SS, Miljko S, et al. Vagus nerve stimulation inhibits cytokine production and attenuates disease severity in rheumatoid arthritis. Proc Natl Acad Sci. 2016;113(29):8284–8289. doi:10.1073/pnas.1605635113
131. Sinniger V, Pellissier S, Fauvelle F, et al. A 12-month pilot study outcomes of vagus nerve stimulation in Crohn’s disease. Neurogastroenterol Motil. 2020;32(10):e13911. doi:10.1111/nmo.13911
132. Stakenborg N, Wolthuis AM, Gomez‐Pinilla PJ, et al. Abdominal vagus nerve stimulation as a new therapeutic approach to prevent postoperative ileus. Neurogastroenterol Motil. 2017;29(9):e13075. doi:10.1111/nmo.13075
133. Salama M, Akan A, Mueller MR. Transcutaneous Stimulation of Auricular Branch of the Vagus Nerve Attenuates the Acute Inflammatory Response After Lung Lobectomy. World J Surg. 2020;44(9):3167–3174. doi:10.1007/s00268-020-05543-w
134. Liu S, Wang ZF, Su YS, et al. Somatotopic Organization and Intensity Dependence in Driving Distinct NPY-Expressing Sympathetic Pathways by Electroacupuncture. Neuron. 2020;108(3):436–450.e7. doi:10.1016/j.neuron.2020.07.015
135. Liu S, Wang Z, Su Y, et al. A neuroanatomical basis for electroacupuncture to drive the vagal–adrenal axis. Nature. 2021;598(7882):641–645. doi:10.1038/s41586-021-04001-4
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.