")
Back to Journals » Journal of Inflammation Research » Volume 17
The Multifaceted Protective Role of Nuclear Factor Erythroid 2-Related Factor 2 in Osteoarthritis: Regulation of Oxidative Stress and Inflammation
Authors Sheng W , Yue Y, Qi T, Qin H, Liu P, Wang D, Zeng H, Yu F
Received 21 May 2024
Accepted for publication 5 September 2024
Published 21 September 2024 Volume 2024:17 Pages 6619—6633
DOI https://doi.org/10.2147/JIR.S479186
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Weibei Sheng,1 Yaohang Yue,1 Tiantian Qi,1 Haotian Qin,1 Peng Liu,1 Deli Wang,1 Hui Zeng,2 Fei Yu1
1Department of Bone & Joint Surgery, National & Local Joint Engineering Research Center of Orthopaedic Biomaterials, Shenzhen Key Laboratory of Orthopaedic Diseases and Biomaterials Research, Peking University Shenzhen Hospital, Shenzhen, 518036, People’s Republic of China; 2Shenzhen Second People’s Hospital, The First Affiliated Hospital of Shenzhen University, Shenzhen, 518035, People’s Republic of China
Correspondence: Hui Zeng; Fei Yu, Email [email protected]; [email protected]
Abstract: Osteoarthritis (OA) is a chronic degenerative joint disease characterized by the degradation of joint cartilage, subchondral bone sclerosis, synovitis, and structural changes in the joint. Recent research has highlighted the role of various genes in the pathogenesis and progression of OA, with nuclear factor erythroid 2-related factor 2 (NRF2) emerging as a critical player. NRF2, a vital transcription factor, plays a key role in regulating the OA microenvironment and slowing the disease’s progression. It modulates the expression of several antioxidant enzymes, such as Heme oxygenase-1 (HO-1) and NAD(P)H oxidoreductase 1 (NQO1), among others, which help reduce oxidative stress. Furthermore, NRF2 inhibits the nuclear factor kappa-B (NF-κB) signaling pathway, thereby decreasing inflammation, joint pain, and the breakdown of cartilage extracellular matrix, while also mitigating cell aging and death. This review discusses NRF2’s impact on oxidative stress, inflammation, cell aging, and various cell death modes (such as apoptosis, necroptosis, and ferroptosis) in OA-affected chondrocytes. The role of NRF2 in OA macrophages, and synovial fibroblasts was also discussed. It also covers NRF2’s role in preserving the cartilage extracellular matrix and alleviating joint pain. The purpose of this review is to provide a comprehensive understanding of NRF2’s protective mechanisms in OA, highlighting its potential as a therapeutic target and underscoring its significance in the development of novel treatment strategies for OA.
Keywords: nuclear factor erythroid 2-related factor 2, osteoarthritis, chondrocytes, inflammation, oxidative stress
Introduction
Osteoarthritis (OA) is a widespread chronic degenerative disease that affects over 22% of the global population aged 40 and above, imposing a significant economic burden on both families and society.1,2 It primarily impacts the knees, hips, and ankles, with the joint microenvironment typically characterized by oxidative stress and inflammation. These conditions lead to cartilage destruction, subchondral bone sclerosis, and synovitis.3 Currently, treatment for early to mid-stage OA focuses on conservative methods, such as anti-inflammatory medications and pain relief, due to the absence of drugs capable of reversing OA’s progression. Although various drugs targeting OA pathology, like anakinra and adalimumab for synovitis, have been developed, they often yield unsatisfactory results.4 This may stem from OA’s complex nature involving multiple joint components, where single-target treatments are insufficient. Hence, ongoing research into more effective therapeutic options is essential.
The onset and progression of OA are driven by complex interactions among various cell types, including chondrocytes, synovial fibroblasts, osteoclasts, macrophages and so on. Chondrocytes, the primary cell type in cartilage, maintain the balance of the cartilage matrix. In the microenvironment of OA, chondrocytes undergo pathological phenotypic changes, leading to increased matrix degradation and chondrocyte apoptosis, resulting in cartilage degeneration.5 Synovial fibroblasts, which secrete synovial fluid to lubricate joints in normal conditions. However, there will be a large amount of production of inflammatory cytokines in OA, leading to synovitis and accelerating cartilage degradation.6 Osteoclasts are involved in bone resorption, and during the progression of OA, subchondral bone loss drives increased bone remodeling and the overactivation of osteoclasts, which induces the catabolism of articular cartilage and promotes the development of OA.7,8 Inflammatory Macrophages drive the inflammatory response by secreting inflammatory mediators, which not only accelerate cartilage destruction but also affect the degradation of other joint structures.9 Therefore, regulating multiple cell types, including chondrocytes, synovial fibroblasts, immune cells, and osteocytes, is crucial for inhibiting the progression of OA.
Discovered in 1994, the transcription factor NF-E2 p45-related factor 2 (NRF2), encoded by NFE2L2, belongs to the human CNC basic leucine zipper transcription factor family.10 NRF2 regulates over 250 genes containing enhancer sequences in their promoter regions, known as antioxidant response elements (ARE). These genes contribute to a synergistic enzyme network responsible for various biochemical processes, including biotransformation reactions, antioxidant metabolism, and the metabolism of carbohydrates, lipids, and proteins.11 Through this network, NRF2 coordinates comprehensive responses to diverse stressors, maintaining cellular stability. Kelch-like ECH-associated protein 1 (Keap1), an inhibitor of NRF2,12 also functions as an E3 ubiquitin ligase substrate adapter,13–15 targeting NRF2 for rapid degradation under non-stress conditions. Under oxidative stress, the highly reactive cysteine residues of KEAP1, when modified by electrophilic molecules, prevent the degradation of NRF2, leading to its accumulation and nuclear translocation. This process triggers dimerization with small MaF proteins, inducing the expression of ARE genes.16,17 These genes encode proteins that perform antioxidant, detoxifying, and anti-inflammatory functions, offering broad cellular protection. The NRF2-Keap1 axis plays a vital role in preventing diseases characterized by oxidative stress and inflammation, including metabolic, inflammatory, autoimmune disorders, and diseases affecting various organs and systems.16,18 NRF2 plays an important regulatory role in OA chondrocytes, osteoclasts, Synovial fibroblasts, and macrophages.9,19–21 NRF2 can inhibit the activation of the NF-κB signaling pathway, thereby suppressing inflammation factors and matrix metalloproteinases (MMPs) in OA chondrocytes, synovial fibroblasts, and macrophages. Additionally, NRF2 regulates the expression of antioxidant-related genes, such as HO-1 and NQO1, reducing oxidative stress levels in the OA microenvironment and thereby decreasing synovial inflammation and protecting the cartilage matrix. Furthermore, NRF2 can inhibit the expression of RANKL factors, thereby suppressing osteoclast activity and maintaining bone homeostasis in the OA environment. This review highlights recent advancements in NRF2 research within the context of OA, offering fresh perspectives for developing novel therapeutics to mitigate the disease.
NRF2 Signaling Pathway in Osteoarthritis, Research Progress
The Role of NRF2 in Chondrocytes
Cartilage destruction stands as a significant pathology in OA. Inflammation and oxidative stress impede the synthesis of the extracellular matrix in chondrocytes, promoting cellular aging and death, which includes apoptosis, necroptosis, and ferroptosis, thus compromising joint cartilage integrity and exacerbating joint pain. NRF2 demonstrates efficacy in suppressing inflammation within chondrocytes by interacting with the NF-κB signaling pathway. Additionally, it orchestrates the expression of various antioxidant enzymes such as HO-1 and NQO1, thus mitigating oxidative stress. As shown in Figure 1, anti-inflammatory and antioxidant properties of NRF2 offer protective mechanisms for compromising’ extracellular matrix, inhibiting cellular aging and death. Subsequent sections delve into a comprehensive analysis of NRF2’s effects on chondrocytes.
 |
Figure 1 The mechanism of NRF2 in regulating chondrocytes, osteoclasts, synovial fibroblasts, and macrophages in OA. Abbreviation, OA, Osteoarthritis; NRF2, nuclear factor erythroid 2-related factor 2; DAMPs, Damage-associated molecular patterns; TLRs, Toll-like receptors; HO-1, Heme Oxygenase-1; NQO1, NAD(P)H,quinone oxidoreductase 1; NF-κB, nuclear factor kappa-B; IkBα, Inhibitor of Nuclear Factor kappa-B alpha; Keap1, kelch-like ECH-associated protein 1; ARE, antioxidant response element; sMaF, small MaF; IL-1β, Interleukin-1β; IL-6, Interleukin-6; TNF-α, Tumor Necrosis Factor Alpha; Ub, Ubiquitination; H2O2, hydrogen peroxide; P-P65, Human phosphorylated nuclear transcription factor P65; ROS, Reactive Oxygen Species; Bax, BCL2 Associated X protein; Bcl2, B-cell lymphoma-2; ΔΨm represents Mitochondrial membrane potential; IL-18, interleukin-18; NO, Nitric Oxide; COX2, Cyclooxygenase-2; MMPs, matrix metalloproteinases; ECM, Extracellular matrix; RANKL, Receptor Activator of Nuclear Factor-κ B Ligand; NFATc1, Nuclear Factor Of Activated T-Cells, Cytoplasmic 1. |
Inflammation Inhibition
The inflammatory response plays a pivotal role in OA pathogenesis. The NF-κB signaling pathway emerges as a crucial player in OA inflammation.22 Stimulated by Interleukin-1β (IL-1β), IκB kinase (IKK) is activated through a series of membrane-proximal events. The phosphorylated IκBs subsequently induce the release of NF-κB, leading to its nuclear translocation and activation of gene transcription, ultimately triggering inflammatory responses.23 This process impedes collagen and proteoglycan production in chondrocytes and activates MMPs and A Disintegrin and Metalloproteinase with Thrombospondin Motifs (ADAMTS), ultimately fostering cartilage degradation.
Augmenting NRF2 nuclear translocation effectively suppresses the NF-κB pathway, thereby mitigating inflammation within chondrocytes and safeguarding joint integrity.24–27 As shown in Table 1, Itaconate,28 Oxymatrine,29 Phillygenin,30 Orientin,31 Stevioside,32 Suramin,33 Chrysophanol,34 Tangeretin,35 Chicoric acid,36 Linalool,37 Ginkgolide C,38 Asiaticoside,39 Corynoline,40 Rhoifolin,41 Procyanidin B2,42 Maltol,43 Lycopene,44 Betulin,45 Limonin,46 Xanthohumol,47 18β-Glycyrrhetinic acid,48 moracin,49 Nomilin,50 hesperetin,51 Akebia Saponin D,52 Sinomenine,53 Sinapic acid,54 Monascin,55 Sauchinone56 and Piceatannol57 have been shown to have the ability to enhance the movement of the NRF2 into the cell nucleus, which in turn inhibits the NF-κB pathway. The mechanism by which NRF2 inhibits the NF-κB signaling pathway can be divided into the following parts. NRF2 suppresses NF-κB activation by increasing the expression of antioxidant enzymes such as HO-1 and NQO1. Additionally, NRF2 indirectly inhibits the nuclear translocation of phosphorylated p65 by modulating the intracellular environment and reducing the level of phosphorylated p65. Furthermore, NRF2 can compete with NF-κB for binding to certain transcription factors and nuclear receptors, thereby reducing NF-κB activity. This inhibition subsequently suppresses downstream pro-inflammatory factors and MMPs while concurrently enhancing the expression of Collagen II and Aggrecan, ultimately preserving cartilage.
 |
Table 1 Activation of NRF2 Inhibits Inflammation and Catabolism in Chondrocytes by Suppressing the NF-κB Signaling Pathway |
Oxidative Stress
Oxidative stress represents a pivotal factor driving age-associated diseases, including OA. An imbalance between Reactive Oxygen Species (ROS) production and the antioxidant capacity of joint cells, such as chondrocytes, constitutes a significant component of OA progression.58 Consequently, alleviating oxidative stress within cartilage can substantially attenuate OA progression. Activation of the NRF2/ARE signaling pathway manifests protective effects against OA pathogenesis by upregulating antioxidant factors such as HO-1, NQO1, Glutathione (GSH), Glutathione Peroxidase (GPx), and Superoxide Dismutase (SOD), thereby suppressing oxidative stress in chondrocytes.59 As shown in Table 2, Fibroblast growth factor 9,60 Curcumin,61 catalase,61 Ellagic acid,62 Allicin,63 Sulforaphane,63 Lycopene63 and Cudratricusxanthone O64 can activate NRF2 in chondrocytes, thereby activating antioxidant enzymes such as HO-1, SOD, and GPx, effectively reducing oxidative stress levels in OA chondrocytes.
 |
Table 2 The Antioxidant and Anti-Chondrocyte Apoptotic Effects of NRF2 |
Regulation of Cartilage Matrix Synthesis
Notably, Sex-determining Region Y (SRY)-box 9 (SOX9) serves as an indispensable transcription factor for chondrocyte lineage differentiation during embryonic development and postnatally in the growth plate and articular chondrocytes.65 Additionally, SOX9 acts as a major driver behind osmolarity-determined chondrogenic differentiation capacity of progenitor cells,66 wherein osmolarity enhances cartilage Extracellular Matrix (ECM) marker expression while specifically affecting ADAMTS4 and ADAMTS5.67 One pivotal function of NRF2 could be to maintain sufficiently high SOX9 expression in articular cartilage throughout aging, thereby mediating ADAMTS suppression to protect cartilage integrity, consequently delaying OA onset.68
Senescence
Age governs NRF2 homeostasis in human articular chondrocytes, with NRF2 protein levels notably lower in older adult chondrocytes (approximately 0.59 fold; P = 0.034) and OA chondrocytes compared to younger cells.69 In OA cartilage, oxidative stress presence upregulates aging-related factors such as Tumor Protein p53 (p53) and Cyclin-Dependent Kinase Inhibitor 2A (p16INK4a), promoting senescence in chondrocytes. The NRF2 signaling pathway regulates the expression of various antioxidant enzymes to inhibit chondrocyte aging. As shown in Table 3, Theaflavin,70 Itaconate,28 Procyanidin B2,42 and S-allyl cysteine71 effectively activate the NRF2 signaling pathway, thereby inhibiting aging in OA chondrocytes.
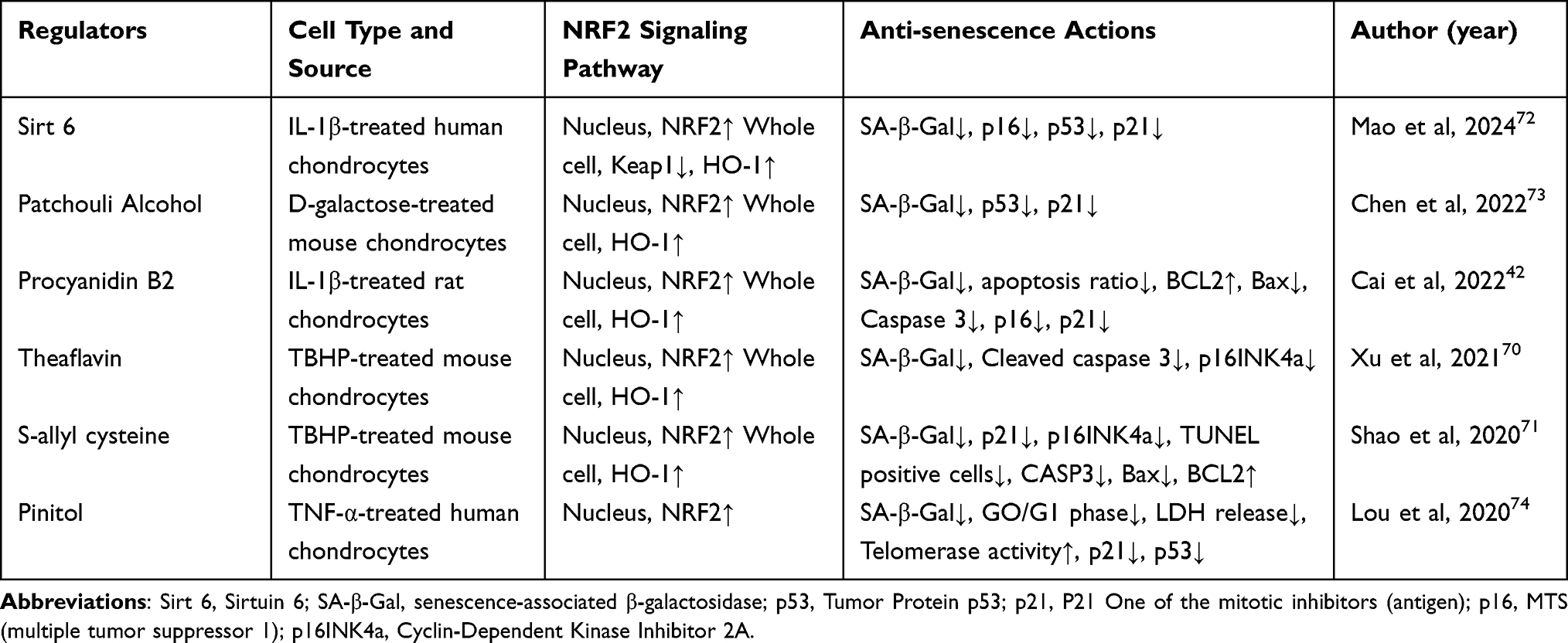 |
Table 3 The Role of NRF2 in Regulating Chondrocyte Senescence |
Apoptosis
Inflammatory environments (such as high IL-1β) and oxidative stress can induce apoptosis in chondrocytes.75,76 NRF2 activation in OA chondrocytes can counteract these effects by inhibiting IL-1β-induced mitochondrial dysfunction, Reactive Oxygen Species (ROS) production, and apoptosis.19 Overexpression of NRF2 upregulates the expression of anti-apoptotic factors, downregulates pro-apoptotic proteins, and activates Extracellular Signal-Regulated Kinase 1/2 (ERK1/2) and its downstream factors, such as ETS-Like Transcription Factor 1 (ELK1), Ribosomal Protein S6 Kinase, 70 kDa (P70S6K), and 90 kDa Ribosomal S6 Kinase (P90RSK).19 Moreover, NRF2 indirectly impacts chondrocyte apoptosis and senescence by controlling the expression of glyoxalase I, an enzyme responsible for detoxifying methylglyoxal.77
Pyroptosis
Pyroptosis is a type of proinflammatory programmed cell death that is triggered by inflammasomes, which is strongly correlated with OA progression.78 It is also reported that activation of the NRF2 signaling pathway may alleviate the progression of OA by suppressing the NOD-like Receptor Protein 3 (NLRP3) inflammasome in primary mouse chondrocytes.79 As shown in Table 4, Cardamonin,80 Ginkgolide C,81 Licochalcone A,79 Loratadine,82 Bisdemethoxycurcumin,83 Cucurbitacin B84 effectively inhibit pyroptosis in OA chondrocytes by activating the NRF2 signaling pathway.
 |
Table 4 Activation of NRF2 Inhibit Chondrocyte Pyroptosis |
Ferroptosis
Ferroptosis, characterized by excessive lipid peroxidation and iron accumulation, is a nonapoptotic cell death process that plays a significant role in the progression of OA.85 The NRF2-ARE system can inhibit or repair lipid peroxidation damage through multiple pathways, thus reducing chondrocytes ferroptosis.86,87 Firstly, key synthesizing enzyme genes of the GSH-GPx4 pathway are positively regulated by NRF2, such as enzymes promoting GSH biosynthesis (glutamate-cysteine ligase, GSH synthetase, and Solute Carrier Family 7 Member A11 (SLC7A11)), GSH reductase, and GPx4.85 NRF2 can also activate the thioredoxin system to compensate for the GSH system.88 Moreover, NRF2 is a central control factor for the expression of NQO1 under steady and stress conditions.89 NQO1, a homodimeric flavoenzyme, can catalyze the reduction of quinones to hydroquinones in a single-step, two-electron reduction reaction; it also plays a significant role in protecting endogenous antioxidants by maintaining the reduced forms of ubiquinone and α-tocopheryl quinone. NRF2 can also regulate the detoxification of lipid peroxidation downstream products, such as by transcriptionally activating the expression of the aldo-keto reductase family (AKR1C1-3) and the aldehyde dehydrogenase family (ALDH3A1).86,87 Finally, NRF2 promotes the expression of ferritin and Ferroportin 1 to store or export free iron, thus reducing intracellular iron accumulation and preventing the occurrence of ferroptosis.90 Targeting NRF2 activation could effectively inhibit ferroptosis in chondrocytes. As shown in Table 5, Gamma-oryzanol,91 Baicalein,92 Curcumin,93 and Deferoxamine94 effectively inhibit cartilage cell ferroptosis by activating the NRF2 signaling pathway.
 |
Table 5 Activation of NRF2 Inhibits Chondrocyte Ferroptosis |
The Role of NRF2 in Macrophages
Macrophages are primarily classified into M1 and M2 types, where M1 macrophages are pro-inflammatory and M2 macrophages are anti-inflammatory.95 In the OA microenvironment, synovial macrophages predominantly differentiate into the M1 type, secreting large amounts of inflammatory cytokines, thereby damaging cartilage and exacerbating the progression of OA. In contrast, M2 macrophages can promote the repair of cartilage.96 Thus, effectively regulating macrophage polarization is a crucial strategy for alleviating OA. Notably, NRF2 can effectively inhibit the differentiation of macrophages into the M1 type and promote differentiation into the M2 type.9 STUB1, also known as CHIP, is a chaperone-dependent E3 ubiquitin ligase that can ubiquitinate NRF2, thus inhibiting its function.97 The research team led by Zheng Wang demonstrated that silencing STUB1 reduces NRF2 ubiquitination, thereby promoting macrophage differentiation into the M2 type and inhibiting the progression of OA. Another research team showed that TRPV1-evoked Ca(2+) influx promoted the phosphorylation of calcium/calmodulin-dependent protein kinase II (CaMKII) and facilitated the nuclear localization of NRF2, ultimately resulting in the inhibition of M1 macrophage polarization.98 These results prove that NRF2 is a key target for regulating macrophage polarization. Regarding the mechanism, NRF2 may inhibit M1 macrophage differentiation by promoting the expression of HO-1, thereby inhibiting the NF-κB signaling pathway. NRF2 may promote M2 macrophage differentiation through the Transforming Growth Factor-beta/SMAD (TGF-β/SMAD) and Janus Kinase/Signal Transducer and Activator of Transcription (JAK/STAT) signaling pathways.9 Besides regulating macrophage polarization, NRF2 can also directly block the transcription of pro-inflammatory cytokines, thereby inhibiting the inflammatory response of macrophages.99 In summary, the activation of NRF2 can inhibit inflammatory macrophages, promote the differentiation of macrophages into a reparative type, and thus effectively protect cartilage and inhibit the progression of OA.
The Role of NRF2 in Synovial Fibroblasts
In the inflammatory microenvironment of OA, macrophages and synovial fibroblasts in the synovium are often in an activated inflammatory state. Activated synovial fibroblasts produce a large amount of inflammatory cytokines (such as IL-1β, Tumor Necrosis Factor-α (TNF-α)), metabolic degradation factors (such as MMPs and ADAMTS), and ROS. These factors further activate the synovial inflammation and damage cartilage. Therefore, inhibiting synovial inflammation is crucial for alleviating OA. The activation of NRF2 is vital for inhibiting the activation of inflammatory synovial fibroblasts. NRF2 activators such as oltipraz can inhibit the hyperactivation of human fibroblasts.21 Carnosine can activate NRF2 and HO-1 expression, effectively inhibiting MMPs and ROS levels in inflammatory synovial fibroblasts and protecting the mitochondrial membrane potential.100 The dihydroartemisinin derivative DC32 can also effectively activates the NRF2 signaling pathway, and concurrently inhibits the NF-κB signaling pathway, thereby effectively suppressing synovial inflammation.101 These results indicate that NRF2 is a key target for inhibiting synovial inflammation, thus protecting cartilage.
Inhibition of Osteoclastogenesis
Osteoclasts impact the development and progression of OA through various mechanisms.7,8 In OA, increased osteoclast activity leads to bone loss, particularly in the subchondral bone, which in turn results in subchondral bone sclerosis and osteophyte formation. Additionally, osteoclasts are activated in the inflammatory environment of arthritis and can secrete inflammatory factors and enzymes, which promote local inflammation. Therefore, inhibiting osteoclast activity is crucial for slowing the progression of OA. The activation of the NRF2/HO-1 signaling pathway can effectively inhibit nuclear factor-κB ligand (RANKL)-induced osteoclast formation and extracellular matrix (ECM) degradation.102
Pain
Pain is a prominent symptom of OA.103 The types of pain associated with OA are still debated. Nerve damage, inflammation, and damaged joint tissues might be the causes of OA pain.104 Pharmacologic treatment of OA pain relies primarily on Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) and opioids.103 NSAIDs are not effective in alleviating OA pain, and opioids have multiple side effects, such as nausea and dizziness.105 Therefore, new treatment options are needed. The activation of NRF2 can alleviate pain behaviors in rats with OA. The activation of NRF2 nuclear transcription can enhance the synthesis of peroxidase enzymes, such as GSH, NQO1, and glutathione S-transferase (GST), leading to a subsequent reduction in the initial pain experienced in OA.106–108
Antioxidants Combined with Nanospheres
NRF2 activators such as Oltipraz, curcumin, and resveratrol suffer from poor water solubility, which significantly impacts their biological activity. With advancements in nanotechnology, issues such as poor drug solubility and rapid degradation can be effectively addressed. Hengfeng Yuan’s team encapsulated the NRF2 activator Oltipraz in ROS-responsive nanoparticles, which, compared to standalone nanoparticles, could effectively activate the NRF2/HO-1 signaling pathway, thereby exhibiting ROS scavenging and anti-apoptotic properties in chondrocytes.109 A bioactive gel based on gallen gum (GG-CD@ARC) encapsulated with the antioxidant arctiin was developed to alleviate the progression of OA by effectively activating NRF2.110 Another study demonstrated that antioxidant arbutin-loaded gelatin methacryloyl-liposome (GM-Lipo@ARB) microspheres were developed to activate the NRF2 signaling pathway, reduce oxidative stress in OA cartilage, and thus alleviate OA.111 Therefore, combining antioxidants with nanotechnology to more efficiently activate the NRF2 signaling pathway could more effectively inhibit the progression of OA.
Conclusion and Prospects
As a transcription factor, NRF2 can affect OA chondrocytes, macrophages, synovial fibroblasts, and osteoclasts. NRF2 inhibits inflammation in OA chondrocytes by suppressing the NF-κB signaling pathway, promotes the expression of various antioxidant enzymes such as HO-1 and NQO1, thus inhibiting oxidative stress, protecting the cartilage extracellular matrix, reducing aging and death of chondrocytes (including apoptosis, ferroptosis, and pyroptosis), and alleviating joint pain. Additionally, NRF2 can inhibit the differentiation of synovial macrophages into the M1 type and promote differentiation into the M2 type, thus creating an environment conducive to cartilage repair. The activation of NRF2 can also inhibit inflammatory synovial fibroblasts, thereby reducing their secretion of pro-inflammatory cytokines and metabolic degradation factors, and protecting the cartilage. NRF2 also inhibits the formation of osteoclasts, thus maintaining the morphology of the subchondral bone. Various drugs have proven effective in alleviating the progression of OA by activating NRF2. Furthermore, the development of nanotechnology can be well integrated with NRF2 activators, activating NRF2 through prolonged drug release, thereby effectively alleviating OA.
Given the protective role of NRF2, researchers should actively explore compounds or drugs that effectively activate NRF2. The specific molecular mechanisms of NRF2 in OA, including its regulatory effects on chondrocytes, synovial fibroblasts, osteoclasts, and macrophages, require further in-depth research. These studies will help to elucidate how NRF2 influences the progression of OA through its antioxidant and anti-inflammatory pathways. Additionally, utilizing gene editing technologies, such as CRISPR-Cas9, to directly regulate NRF2 gene expression has emerged as a novel therapeutic strategy.
While NRF2 exhibits various benefits, current research on NRF2 still faces several challenges and difficulties. Most of the NRF2 activators developed so far lack high selectivity, and while activating the antioxidant pathway, they may also activate other signaling pathways, such as glycolysis and mitochondrial function, leading to unintended biological effects.112,113 Furthermore, although NRF2 activators may show positive effects in the short term, the safety and side effects of long-term use still require further investigation. Additionally, while nanocarriers can enhance the efficacy of NRF2 activators, their drug-loading efficiency and sustained-release properties need further improvement. Finally, although NRF2 shows great potential in laboratory studies, translating this potential into clinical applications remains challenging. Currently, clinical research on NRF2 activators in OA remains in its early stages. Most studies are focused on preclinical models, and there are still relatively few clinical trials specifically targeting OA with NRF2 activators. The safety and efficacy of these compounds in long-term human use are still under investigation.
In summary, activation of NRF2 can effectively regulate the function of chondrocytes, synovial macrophages, synovial fibroblasts, and osteoclasts, thereby effectively inhibiting synovial inflammation, protecting cartilage and subchondral bone, alleviating joint pain, and serving as a potential target for treating OA. However, researchers still need to overcome issues of selectivity and safety, as well as challenges related to drug delivery, to achieve clinical application of NRF2 activators. Addressing these challenges will open new prospects for the treatment of OA.
Acknowledgments
This study was supported by grants from Guangdong Basic and Applied Basic Research Foundation (No. 2021A1515220037, 2022A1515220165), National Natural Science Foundation of China (No. 82172432), Shenzhen Science and Technology Program (No. JCYJ20210324110214040), Shenzhen Key Laboratory of Orthopaedic Diseases and Biomaterials Research (No. ZDSYS20220606100602005) and Sanming Project of Medicine in Shenzhen (No. SZSM202211038).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Mao X, Yan B, Chen H, Lai P, Ma J. BRG1 mediates protective ability of spermidine to ameliorate osteoarthritic cartilage by Nrf2/KEAP1 and STAT3 signaling pathway. Int immunopharmacol. 2023;122:110593. doi:10.1016/j.intimp.2023.110593
2. Pigeolet M, Jayaram A, Park KB, Meara JG. Osteoarthritis in 2020 and beyond. Lancet. 2021;397(10279):1059–1060. doi:10.1016/S0140-6736(21)00208-7
3. Katz JN, Arant KR, Loeser RF. Diagnosis and treatment of hip and knee osteoarthritis. A Review, Jama. 2021;325(6):568–578. doi:10.1001/jama.2020.22171
4. Latourte A, Kloppenburg M, Richette P. Emerging pharmaceutical therapies for osteoarthritis, Nature reviews. Rheumatology. 2020;16:673–688. doi:10.1038/s41584-020-00518-6
5. Charlier E, Deroyer C, Ciregia F, et al. Chondrocyte dedifferentiation and osteoarthritis (OA. Biochem Pharmacol. 2019;165:49–65. doi:10.1016/j.bcp.2019.02.036
6. Chen HW, Huang CH, Huang CF, Chang CH, Liao HJ. Distinct subsets of synovial fibroblasts control cartilage destruction in joint diseases. Cli Experim rheumato. 2024;42(5):1118–1126. doi:10.55563/clinexprheumatol/txl9rm
7. Wang H, Yuan T, Wang Y, et al. Osteoclasts and osteoarthritis, novel intervention targets and therapeutic potentials during aging. Aging Cell. 2024;23:e14092. doi:10.1111/acel.14092
8. Burr DB, Gallant MA. Bone remodelling in osteoarthritis, Nature reviews. Rheumatology. 2012;8:665–673. doi:10.1038/nrrheum.2012.130
9. Wang L, He C. Nrf2-mediated anti-inflammatory polarization of macrophages as therapeutic targets for osteoarthritis. Front Immunol. 2022;13:967193.
10. Moi P, Chan K, Asunis I, Cao A, Kan YW. Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic leucine zipper transcriptional activator that binds to the tandem NF-E2/AP1 repeat of the beta-globin locus control region. Proc Natl Acad Sci USA. 1994;91(21):9926–9930. doi:10.1073/pnas.91.21.9926
11. Hayes JD, Dinkova-Kostova AT. The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem Sci. 2014;39(4):199–218. doi:10.1016/j.tibs.2014.02.002
12. Itoh K, Wakabayashi N, Katoh Y, et al. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999;13(1):76–86. doi:10.1101/gad.13.1.76
13. Zhang DD, Lo SC, Cross JV, Templeton DJ, Hannink M. Keap1 is a redox-regulated substrate adaptor protein for a Cul3-dependent ubiquitin ligase complex. Mol Cell Biol. 2004;24(24):10941–10953. doi:10.1128/MCB.24.24.10941-10953.2004
14. Cullinan SB, Gordan JD, Jin J, Harper JW, Diehl JA. The Keap1-BTB protein is an adaptor that bridges Nrf2 to a Cul3-based E3 ligase, oxidative stress sensing by a Cul3-Keap1 ligase. Mol Cell Biol. 2004;24(19):8477–8486. doi:10.1128/MCB.24.19.8477-8486.2004
15. Kobayashi A, Kang MI, Okawa H, et al. Oxidative stress sensor Keap1 functions as an adaptor for Cul3-based E3 ligase to regulate proteasomal degradation of Nrf2. Mol Cell Biol. 2004;24(16):7130–7139. doi:10.1128/MCB.24.16.7130-7139.2004
16. Cuadrado A, Manda G, Hassan A, et al. transcription factor NRF2 as a therapeutic target for chronic diseases, a systems medicine approach. Pharmacol Rev. 2018;70(2):348–383. doi:10.1124/pr.117.014753
17. Yamamoto M, Kensler TW, Motohashi H. The KEAP1-NRF2 System, a thiol-based sensor-effector apparatus for maintaining redox homeostasis. Pharmacol Rev. 2018;98(3):1169–1203. doi:10.1152/physrev.00023.2017
18. Liby KT, Sporn MB. Synthetic oleanane triterpenoids, multifunctional drugs with a broad range of applications for prevention and treatment of chronic disease. Pharmacol Rev. 2012;64(4):972–1003. doi:10.1124/pr.111.004846
19. Khan NM, Ahmad I, Haqqi TM. Nrf2/ARE pathway attenuates oxidative and apoptotic response in human osteoarthritis chondrocytes by activating ERK1/2/ELK1-P70S6K-P90RSK signaling axis. Free Radic Biol Med. 2018;116:159–171. doi:10.1016/j.freeradbiomed.2018.01.013
20. Hyeon S, Lee H, Yang Y, Jeong W. Nrf2 deficiency induces oxidative stress and promotes RANKL-induced osteoclast differentiation. Free Radic Biol Med. 2013;65:789–799. doi:10.1016/j.freeradbiomed.2013.08.005
21. Jie P, Wu Y, Song C, Cheng Y, Liu Y, Chen K. Mechanism of Nrf2/miR338-3p/TRAP-1 pathway involved in hyperactivation of synovial fibroblasts in patients with osteoarthritis. Heliyon. 2023;9(11):e21412. doi:10.1016/j.heliyon.2023.e21412
22. Scanzello CR. Role of low-grade inflammation in osteoarthritis. Curr opinion Rheumatol. 2017;29(1):79–85. doi:10.1097/BOR.0000000000000353
23. Rigoglou S, Papavassiliou AG. The NF-κB signalling pathway in osteoarthritis. Int J. Biochem Cell Biol. 2013;45(11):2580–2584. doi:10.1016/j.biocel.2013.08.018
24. Abusarah J, Benabdoune H, Shi Q, et al. Elucidating the role of protandim and 6-gingerol in protection against osteoarthritis. J Cell Biochem. 2017;118:1003–1013. doi:10.1002/jcb.25659
25. Khan NM, Haseeb A, Ansari MY, Devarapalli P, Haynie S, Haqqi TM. Wogonin, a plant derived small molecule, exerts potent anti-inflammatory and chondroprotective effects through the activation of ROS/ERK/Nrf2 signaling pathways in human Osteoarthritis chondrocytes. Free Radic Biol Med. 2017;106:288–301. doi:10.1016/j.freeradbiomed.2017.02.041
26. Wang Y, Chen Y, Chen Y, Zhou B, Shan X, Yang G. Eriodictyol inhibits IL-1β-induced inflammatory response in human osteoarthritis chondrocytes. Biomed pharmaco. 2018;107:1128–1134. doi:10.1016/j.biopha.2018.08.103
27. Liang J, Wang S, Hu J, et al. Targeted inhibition of TXNRD1 prevents cartilage extracellular matrix degeneration by activating Nrf2 pathway in osteoarthritis. Biochem Biophy Rese Commun. 2022;635:267–276. doi:10.1016/j.bbrc.2022.10.059
28. Ni L, Lin Z, Hu S, et al. Itaconate attenuates osteoarthritis by inhibiting STING/NF-κB axis in chondrocytes and promoting M2 polarization in macrophages. Biochem Pharmacol. 2022;198:114935. doi:10.1016/j.bcp.2022.114935
29. Zhou K, Liu D, Jin Y, Xia W, Zhang P, Zhou Z. Oxymatrine ameliorates osteoarthritis via the Nrf2/NF-κB axis in vitro and in vivo. Chem Biol Interact. 2023;380:110539. doi:10.1016/j.cbi.2023.110539
30. Zhang P, Jin Y, Xia W, Wang X, Zhou Z. Phillygenin inhibits inflammation in chondrocytes via the Nrf2/NF-κB axis and ameliorates osteoarthritis in mice. J Orthop Transl. 2023;41:1–11. doi:10.1016/j.jot.2023.03.002
31. Xia W, Xiao J, Tong C, et al. Orientin inhibits inflammation in chondrocytes and attenuates osteoarthritis through Nrf2/NF-κB and SIRT6/NF-κB pathway. J Ortho Res. 2023;41(11):2405–2417. doi:10.1002/jor.25573
32. Wu J, Li H, Hu F, Luo P. Stevioside attenuates osteoarthritis via regulating Nrf2/HO-1/NF-κB pathway. J Orthop Transl. 2023;38:190–202. doi:10.1016/j.jot.2022.05.005
33. Shen PC, Huang SH, Liu ZM, Lu CC, Chou SH, Tien Y. Tien, suramin ameliorates osteoarthritis by acting on the Nrf2/HO-1 and NF-κB signaling pathways in chondrocytes and promoting M2 polarization in macrophages. Int Immunopharmacol. 2023;120:110295. doi:10.1016/j.intimp.2023.110295
34. Lu J, Miao Z, Jiang Y, et al. Chrysophanol prevents IL-1β-Induced inflammation and ECM degradation in osteoarthritis via the Sirt6/NF-κB and Nrf2/NF-κB axis. Biochem Pharmaco. 2023;208:115402. doi:10.1016/j.bcp.2022.115402
35. Shi Y, Chen J, Li S, et al. Tangeretin suppresses osteoarthritis progression via the Nrf2/NF-κB and MAPK/NF-κB signaling pathways. Phytomed, Int J Phyt Phytop. 2022;98:153928. doi:10.1016/j.phymed.2022.153928
36. Qu Y, Shen Y, Teng L, et al. Chicoric acid attenuates tumor necrosis factor-α-induced inflammation and apoptosis via the Nrf2/HO-1, PI3K/AKT and NF-κB signaling pathways in C28/I2 cells and ameliorates the progression of osteoarthritis in a rat model. Int Immunopharmacol. 2022;111:109129. doi:10.1016/j.intimp.2022.109129
37. Miao Z, Dong M, Wang Z, Ma J, Lin Y, Wu Y. Linalool inhibits the progression of osteoarthritis via the Nrf2/HO-1 signal pathway both in vitro and in vivo. Int immunopharmacol. 2022;113:109338. doi:10.1016/j.intimp.2022.109338
38. Ma T, Jia L, Zhao J, et al. Ginkgolide C slows the progression of osteoarthritis by activating Nrf2/HO-1 and blocking the NF-κB pathway. Front Pharmacol. 2022;13:1027553. doi:10.3389/fphar.2022.1027553
39. Luo P, Huang Q, Chen S, Wang Y, Dou H. Asiaticoside ameliorates osteoarthritis progression through activation of Nrf2/HO-1 and inhibition of the NF-κB pathway. Int immunopharmacol. 2022;108:108864. doi:10.1016/j.intimp.2022.108864
40. Li S, Shi Y, Zhang S, et al. Corynoline alleviates osteoarthritis development via the Nrf2/NF-κB pathway. Oxi Med Cell Longe. 2022;2022:2188145. doi:10.1155/2022/2188145
41. Chen H, Qin J, Shi H, Li Q, Zhou S, Chen L. Rhoifolin ameliorates osteoarthritis via the Nrf2/NF-κB axis, in vitro and in vivo experiments. Osteoarth Cartila. 2022;30:735–745. doi:10.1016/j.joca.2022.01.009
42. Cai W, Zhang Y, Jin W, et al. Procyanidin B2 ameliorates the progression of osteoarthritis, An in vitro and in vivo study. Int Immunopharmacol. 2022;113:109336. doi:10.1016/j.intimp.2022.109336
43. Zhu DC, Wang YH, Lin JH, Miao ZM, Xu JJ, Wu YS. Maltol inhibits the progression of osteoarthritis via the nuclear factor-erythroid 2–related factor-2/heme oxygenase-1 signal pathway in vitro and in vivo. Food Funct. 2021;12(3):1327–1337. doi:10.1039/D0FO02325F
44. Zhan J, Yan Z, Kong X, et al. Lycopene inhibits IL-1β-induced inflammation in mouse chondrocytes and mediates murine osteoarthritis. J Cell & Mol Med. 2021;25(7):3573–3584. doi:10.1111/jcmm.16443
45. Ren C, Jin J, Hu W, et al. Betulin alleviates the inflammatory response in mouse chondrocytes and ameliorates osteoarthritis via AKT/Nrf2/HO-1/NF-κB Axis. Front in Pharmacolo. 2021;12:754038. doi:10.3389/fphar.2021.754038
46. Jin J, Lv X, Wang B, et al. Limonin inhibits IL-1 β -induced inflammation and catabolism in chondrocytes and ameliorates osteoarthritis by activating Nrf2. Oxida Med Cell Lon. 2021;2021(1):7292512. doi:10.1155/2021/7292512
47. Chen X, Li Z, Hong H, et al. Xanthohumol suppresses inflammation in chondrocytes and ameliorates osteoarthritis in mice. Biomed Pharmaco. 2021;137:111238. doi:10.1016/j.biopha.2021.111238
48. Chen B, Zhu D, Xie C, et al. 18β-Glycyrrhetinic acid inhibits IL-1β-induced inflammatory response in mouse chondrocytes and prevents osteoarthritic progression by activating Nrf2. Food Funct. 2021;12(18):8399–8410. doi:10.1039/D1FO01379C
49. Zhou S, Shi J, Wen H, Xie W, Han X, Li H. A chondroprotective effect of moracin on IL-1β-induced primary rat chondrocytes and an osteoarthritis rat model through Nrf2/HO-1 and NF-κB axes. Food Funct. 2020;11(9):7935–7945. doi:10.1039/D0FO01496F
50. Xue XH, Xue JX, Hu W, Shi FL, Yang Y. Nomilin targets the Keap1-Nrf2 signalling and ameliorates the development of osteoarthritis. J Cell & Mol Med. 2020;24(15):8579–8588. doi:10.1111/jcmm.15484
51. Lin Z, Fu C, Yan Z, et al. The protective effect of hesperetin in osteoarthritis: an in vitro and in vivo study. Food funct. 2020;11(3):2654–2666. doi:10.1039/C9FO02552A
52. Gu M, Jin J, Ren C, et al. Akebia Saponin D suppresses inflammation in chondrocytes via the NRF2/HO-1/NF-κB axis and ameliorates osteoarthritis in mice. Food Funct. 2020;11(12):10852–10863. doi:10.1039/D0FO01909G
53. Wu Y, Lin Z, Yan Z, Wang Z, Fu X, Yu K. Sinomenine contributes to the inhibition of the inflammatory response and the improvement of osteoarthritis in mouse-cartilage cells by acting on the Nrf2/HO-1 and NF-κB signaling pathways. Int Immunopharmacol. 2019;75:105715. doi:10.1016/j.intimp.2019.105715
54. Li X, Lin J, Ding X, et al. The protective effect of sinapic acid in osteoarthritis, in vitro and in vivo studies. J Cell & Mol Med. 2019;23(3):1940–1950. doi:10.1111/jcmm.14096
55. Zheng G, Zhan Y, Tang Q, et al. Monascin inhibits IL-1β induced catabolism in mouse chondrocytes and ameliorates murine osteoarthritis. Food funct. 2018;9(3):1454–1464. doi:10.1039/C7FO01892D
56. Wu D, Jin S, Lin Z, et al. Sauchinone inhibits IL-1β induced catabolism and hypertrophy in mouse chondrocytes to attenuate osteoarthritis via Nrf2/HO-1 and NF-κB pathways. Int Immunopharmacol. 2018;62:181–190. doi:10.1016/j.intimp.2018.06.041
57. Tang Q, Feng Z, Tong M, et al. Piceatannol inhibits the IL-1β-induced inflammatory response in human osteoarthritic chondrocytes and ameliorates osteoarthritis in mice by activating Nrf2. Food Funct. 2017;8:3926–3937. doi:10.1039/C7FO00822H
58. Bolduc JA, Collins JA, Loeser RF. Reactive oxygen species, aging and articular cartilage homeostasis. Free Rad Biol Med. 2019;132:73–82. doi:10.1016/j.freeradbiomed.2018.08.038
59. Ashrafizadeh M, Fekri HS, Ahmadi Z, Farkhondeh T, Samarghandian S. Therapeutic and biological activities of berberine, The involvement of Nrf2 signaling pathway. J Cell Biochem. 2020;121(2):1575–1585. doi:10.1002/jcb.29392
60. Pan YN, Jia C, Yu JP, Wu ZW, Xu GC, Huang YX. Fibroblast growth factor 9 reduces TBHP-induced oxidative stress in chondrocytes and diminishes mouse osteoarthritis by activating ERK/Nrf2 signaling pathway. Int Immunopharmacol. 2023;114:109606. doi:10.1016/j.intimp.2022.109606
61. Chen B, He Q, Chen C, et al. Combination of curcumin and catalase protects against chondrocyte injury and knee osteoarthritis progression by suppressing oxidative stress. Biomed Pharmacother. 2023;168:115751. doi:10.1016/j.biopha.2023.115751
62. Zhu W, Tang H, Li J, Guedes RM, Cao L, Guo C. Ellagic acid attenuates interleukin-1β-induced oxidative stress and exerts protective effects on chondrocytes through the Kelch-like ECH-associated protein 1 (Keap1)/ Nuclear factor erythroid 2-related factor 2 (Nrf2) pathway. Bioengineered. 2022;13(4):9233–9247. doi:10.1080/21655979.2022.2059995
63. Yang J, Song X, Feng Y, et al. Natural ingredients-derived antioxidants attenuate H(2)O(2)-induced oxidative stress and have chondroprotective effects on human osteoarthritic chondrocytes via Keap1/Nrf2 pathway. Free Rad Biol Med. 2020;152:854–864. doi:10.1016/j.freeradbiomed.2020.01.185
64. Kim EN, Lee HS, Jeong GS. Cudratricusxanthone O inhibits H(2)O(2)-induced cell damage by activating Nrf2/HO-1 pathway in human chondrocytes. Antioxidants. 2020;10(1):9. doi:10.3390/antiox10010009
65. Henry SP, Liang S, Akdemir KC, de Crombrugghe B. The postnatal role of Sox9 in cartilage. J Bone Miner Res. 2012;27(12):2511–2525. doi:10.1002/jbmr.1696
66. Caron MM, van der Windt AE, Emans PJ, van Rhijn LW, Jahr H, Welting TJ. Osmolarity determines the in vitro chondrogenic differentiation capacity of progenitor cells via nuclear factor of activated T-cells 5. Bone. 2013;53(1):94–102. doi:10.1016/j.bone.2012.11.032
67. van der Windt AE, Haak E, Das RH, et al. Physiological tonicity improves human chondrogenic marker expression through nuclear factor of activated T-cells 5 in vitro. Arthritis Res Ther. 2010;12(3):R100. doi:10.1186/ar3031
68. Kubo Y, Beckmann R, Fragoulis A, et al. Nrf2/ARE signaling directly regulates SOX9 to potentially alter age-dependent cartilage degeneration. Antioxidants. 2022;12(1):11. doi:10.3390/antiox12010011
69. Taylor EL, Collins JA, Gopalakrishnan P, Chubinskaya S, Loeser RF. Age and oxidative stress regulate Nrf2 homeostasis in human articular chondrocytes. Osteoarthr Cartila. 2023;31(9):1214–1223. doi:10.1016/j.joca.2023.05.004
70. Xu XX, Zheng G, Tang SK, Liu HX, Hu YZ, Shang P. Theaflavin protects chondrocytes against apoptosis and senescence via regulating Nrf2 and ameliorates murine osteoarthritis. Food Funct. 2021;12(4):1590–1602. doi:10.1039/D0FO02038A
71. Shao Z, Pan Z, Lin J, et al. S-allyl cysteine reduces osteoarthritis pathology in the tert-butyl hydroperoxide-treated chondrocytes and the destabilization of the medial meniscus model mice via the Nrf2 signaling pathway. Aging. 2020;12(19):19254–19272. doi:10.18632/aging.103757
72. Mao LW, Jiang QY, Meng N, et al. Sirt6 promotes DNA damage repair in osteoarthritis chondrocytes by activating the Keap1/Nrf2/HO-1 signaling pathway. Cell Cycle. 2024;23:1–13.
73. Chen M, Wen H, Zhou S, Yan X, Li H. Patchouli alcohol inhibits D-Gal induced oxidative stress and ameliorates the quality of aging cartilage via activating the Nrf2/HO-1 pathway in mice. Oxid Med Cell Longev. 2022;2022:6821170. doi:10.1155/2022/6821170
74. Lou C, Deng A, Zheng H, et al. Pinitol suppresses TNF-α-induced chondrocyte senescence. Cytokine. 2020;130:155047. doi:10.1016/j.cyto.2020.155047
75. Schuerwegh AJ, Dombrecht EJ, Stevens WJ, Van Offel JF, Bridts CH, De Clerck LS. Influence of pro-inflammatory (IL-1 alpha, IL-6, TNF-alpha, IFN-gamma) and anti-inflammatory (IL-4) cytokines on chondrocyte function. Osteoarthritis Cartilage. 2003;11(9):681–687. doi:10.1016/S1063-4584(03)00156-0
76. Li D, Ni S, Miao KS, Zhuang C. PI3K/Akt and caspase pathways mediate oxidative stress-induced chondrocyte apoptosis. Cell Stress Chapero. 2019;24(1):195–202. doi:10.1007/s12192-018-0956-4
77. Ahmed U, Thornalley PJ, Rabbani N. Possible role of methylglyoxal and glyoxalase in arthritis. Biochem Soc Trans. 2014;42(2):538–542. doi:10.1042/BST20140024
78. An S, Hu H, Li Y, Hu Y. Pyroptosis plays a role in osteoarthritis. Aging and Disease. 2020;11(5):1146–1157. doi:10.14336/AD.2019.1127
79. Yan Z, Qi W, Zhan J, et al. Activating Nrf2 signalling alleviates osteoarthritis development by inhibiting inflammasome activation. J Cell & Mol Med. 2020;24(22):13046–13057. doi:10.1111/jcmm.15905
80. Jiang J, Cai M. Cardamonin inhibited IL-1β induced injury by inhibition of NLRP3 inflammasome via activating Nrf2/NQO-1 signaling pathway in chondrocyte. J Microbiol Biotechn. 2021;31(6):794–802. doi:10.4014/jmb.2103.03057
81. Jia L, Gong Y, Jiang X, et al. Ginkgolide C inhibits ROS-mediated activation of NLRP3 inflammasome in chondrocytes to ameliorate osteoarthritis. J Ethnopharmacol. 2024;325:117887. doi:10.1016/j.jep.2024.117887
82. Gao F, Zhang S. Loratadine alleviates advanced glycation end product-induced activation of NLRP3 inflammasome in human chondrocytes. Drug Design, Develop Ther. 2020;14:2899–2908. doi:10.2147/DDDT.S243512
83. Jin G, Xu W, Tang H, Cui Y, Zhang H. Bisdemethoxycurcumin, a curcumin, protects chondrocytes, and reduces cartilage inflammation via the NRF2/HO-1/NLRP3 pathway. Immun Inflamm Dis. 2024;12(2):e1195. doi:10.1002/iid3.1195
84. Lou C, Fang Y, Mei Y, et al. Cucurbitacin B attenuates osteoarthritis development by inhibiting NLRP3 inflammasome activation and pyroptosis through activating Nrf2 / HO −1 pathway. Phytotherapy Research, PTR. 2024;38(7):3352–3369. doi:10.1002/ptr.8209
85. Dixon SJ, Lemberg KM, Lamprecht MR, et al. Ferroptosis, an iron-dependent form of nonapoptotic cell death. Cell. 2012;149(5):1060–1072. doi:10.1016/j.cell.2012.03.042
86. Dodson M, Castro-Portuguez R, Zhang DD. NRF2 plays a critical role in mitigating lipid peroxidation and ferroptosis. Redox Biology. 2019;23:101107. doi:10.1016/j.redox.2019.101107
87. Zakharova ET, Sokolov AV, Pavlichenko NN, et al. Erythropoietin and Nrf2, key factors in the neuroprotection provided by apo-lactoferrin. BiometalsInt J Role Metal Ions iBiology, Biochem, Med. 2018;31(3):425–443. doi:10.1007/s10534-018-0111-9
88. Kim YC, Masutani H, Yamaguchi Y, Itoh K, Yamamoto M, Yodoi J. Hemin-induced activation of the thioredoxin gene by Nrf2. A differential regulation of the antioxidant responsive element by a switch of its binding factors. J Biol Chem. 2001;276(21):18399–18406. doi:10.1074/jbc.M100103200
89. Tanigawa S, Fujii M, Hou DX. Action of Nrf2 and Keap1 in ARE-mediated NQO1 expression by quercetin. Free Radic Biol Med. 2007;42(11):1690–1703. doi:10.1016/j.freeradbiomed.2007.02.017
90. Yang X, Park SH, Chang HC, et al. Sirtuin 2 regulates cellular iron homeostasis via deacetylation of transcription factor NRF2. J Clin Investig. 2017;127(4):1505–1516. doi:10.1172/JCI88574
91. Dai ZH, Zhou CC, Yu CY, et al. Gamma-oryzanol alleviates osteoarthritis development by targeting KEAP1-Nrf2 binding to interfere with chondrocyte ferroptosis. Int Immunopharmacol. 2024;128:111469. doi:10.1016/j.intimp.2023.111469
92. Wan Y, Shen K, Yu H, Fan W. Baicalein limits osteoarthritis development by inhibiting chondrocyte ferroptosis. Free Radic Biol Med. 2023;196:108–120. doi:10.1016/j.freeradbiomed.2023.01.006
93. Zhou Y, Jia Z, Wang J, et al. Curcumin reverses erastin-induced chondrocyte ferroptosis by upregulating Nrf2. Heliyon. 2023;9(10):e20163. doi:10.1016/j.heliyon.2023.e20163
94. Guo Z, Lin J, Sun K, et al. Deferoxamine alleviates osteoarthritis by inhibiting chondrocyte ferroptosis and activating the Nrf2 Pathway. Front Pharmacol. 2022;13:791376. doi:10.3389/fphar.2022.791376
95. Chen S, Saeed A, Liu Q, et al. Macrophages in immunoregulation and therapeutics. Signal Transduct Target Ther. 2023;8(1):207. doi:10.1038/s41392-023-01452-1
96. Xie J, Huang Z, Yu X, Zhou L, Pei F. Clinical implications of macrophage dysfunction in the development of osteoarthritis of the knee. Cytokine Growth Factor Rev. 2019;46:36–44. doi:10.1016/j.cytogfr.2019.03.004
97. Cao N, Wang D, Liu B, et al. Silencing of STUB1 relieves osteoarthritis via inducing NRF2-mediated M2 macrophage polarization. Mol Immunol. 2023;164:112–122. doi:10.1016/j.molimm.2023.11.010
98. Lv Z, Xu X, Sun Z, et al. TRPV1 alleviates osteoarthritis by inhibiting M1 macrophage polarization via Ca(2+)/CaMKII/Nrf2 signaling pathway. Cell Death Dise. 2021;12(6):504. doi:10.1038/s41419-021-03792-8
99. Kobayashi EH, Suzuki T, Funayama R, et al. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat Communi. 2016;7(1):11624. doi:10.1038/ncomms11624
100. Busa P, Lee SO, Huang N, Kuthati Y, Wong CS. Carnosine alleviates knee osteoarthritis and promotes synoviocyte protection via activating the Nrf2/HO-1 signaling pathway, an in-vivo and in-vitro study. Antioxidants. 2022;11(6):1209. doi:10.3390/antiox11061209
101. Li YN, Fan ML, Liu HQ, et al. Dihydroartemisinin derivative DC32 inhibits inflammatory response in osteoarthritic synovium through regulating Nrf2/NF-κB pathway. Int Immunopharmacol. 2019;74:105701. doi:10.1016/j.intimp.2019.105701
102. Yang R, Guo Y, Zong S, et al. Bardoxolone methyl ameliorates osteoarthritis by inhibiting osteoclastogenesis and protecting the extracellular matrix against degradation. Heliyon. 2023;9(2):e13080. doi:10.1016/j.heliyon.2023.e13080
103. Bijlsma JW, Berenbaum F, Lafeber FP. Osteoarthritis, an update with relevance for clinical practice. Lancet. 2011;377(9783):2115–2126. doi:10.1016/S0140-6736(11)60243-2
104. O’Neill TW, Felson DT. Mechanisms of Osteoarthritis (OA) Pain. Current Osteop Reports. 2018;16(5):611–616. doi:10.1007/s11914-018-0477-1
105. Schjerning AM, McGettigan P, Gislason G. Cardiovascular effects and safety of (non-aspirin) NSAIDs, Nature reviews. Cardiology. 2020;17(9):574–584. doi:10.1038/s41569-020-0366-z
106. Hsu DZ, Chu PY, Jou IM. Enteral sesame oil therapeutically relieves disease severity in rat experimental osteoarthritis. Food Nutr Res. 2016;60(1):29807. doi:10.3402/fnr.v60.29807
107. Hsu DZ, Chu PY, Jou IM. Daily sesame oil supplement attenuates joint pain by inhibiting muscular oxidative stress in osteoarthritis rat model. J Nutr Biochem. 2016;29:36–40. doi:10.1016/j.jnutbio.2015.10.007
108. Sun J, Wang XH, Song FH, et al. Inhibition of Brd4 alleviates osteoarthritis pain via suppression of neuroinflammation and activation of Nrf2-mediated antioxidant signalling. Brit j Pharmac. 2023;180(24):3194–3214. doi:10.1111/bph.16195
109. Jiang Z, Wang H, Zhang Z, Pan J, Yuan H. Cartilage targeting therapy with reactive oxygen species-responsive nanocarrier for osteoarthritis. J Nanobiotechnology. 2022;20(1):419. doi:10.1186/s12951-022-01629-w
110. Liu Y, Hou M, Pan Z, et al. Arctiin-reinforced antioxidant microcarrier antagonizes osteoarthritis progression. J Nanobiotechnology. 2022;20(1):303. doi:10.1186/s12951-022-01505-7
111. Jin J, Liu Y, Jiang C, et al. Arbutin-modified microspheres prevent osteoarthritis progression by mobilizing local anti-inflammatory and antioxidant responses. Materials Today Bio. 2022;16:100370. doi:10.1016/j.mtbio.2022.100370
112. Mitsuishi Y, Taguchi K, Kawatani Y, et al. Nrf2 redirects glucose and glutamine into anabolic pathways in metabolic reprogramming. Cancer Cell. 2012;22(1):66–79. doi:10.1016/j.ccr.2012.05.016
113. Pang S, Lynn DA, Lo JY, Paek J, Curran SP. SKN-1 and Nrf2 couples proline catabolism with lipid metabolism during nutrient deprivation. Nat Communic. 2014;5(1):5048. doi:10.1038/ncomms6048
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.