")
Back to Journals » Journal of Inflammation Research » Volume 17
The Onset of Systemic Lupus Erythematosus Triggers Nucleus Pulposus Cell Pyroptosis to Exacerbate Intervertebral Disc Degeneration
Authors Lao Z, Fang X, Shen S, Zhang Y, Chen X, Zhang H, Bian Y, Zhou C, Bao R, Xu T , Jin H, Fu F, Wu C, Hu C , Ruan H
Received 10 July 2024
Accepted for publication 18 October 2024
Published 25 October 2024 Volume 2024:17 Pages 7705—7719
DOI https://doi.org/10.2147/JIR.S486297
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Zhaobai Lao,1,* Xuliang Fang,1,* Shuchao Shen,1,* Yuliang Zhang,2,* Xin Chen,1 Helou Zhang,1 Yishan Bian,1 Chengcong Zhou,1 Ronghua Bao,2 Taotao Xu,3 Hongting Jin,1 Fangda Fu,1 Chengliang Wu,1 Changfeng Hu,4 Hongfeng Ruan1
1Institute of Orthopaedics and Traumatology, The First Affiliated Hospital of Zhejiang Chinese Medical University (Zhejiang Provincial Hospital of Traditional Chinese Medicine), Hangzhou, Zhejiang, 310053, People’s Republic of China; 2Hangzhou Fuyang Hospital of TCM Orthopedics and Traumatology, Hangzhou, Zhejiang, 311400, People’s Republic of China; 3Department of Orthopaedics, The First Affiliated Hospital of Zhejiang Chinese Medical University (Zhejiang Provincial Hospital of Traditional Chinese Medicine), Hangzhou, Zhejiang, 310053, People’s Republic of China; 4College of Basic Medical Sciences, Zhejiang Chinese Medical University, Hangzhou, Zhejiang, 310053, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Changfeng Hu, College of Basic Medical Sciences, Zhejiang Chinese Medical University, Hangzhou, Zhejiang, 310053, People’s Republic of China, Email [email protected] Hongfeng Ruan, Institute of Orthopaedics and Traumatology, The First Affiliated Hospital of Zhejiang Chinese Medical University, 548 Binwen Road, Hangzhou, 310053, People’s Republic of China, Email [email protected]
Purpose: Systemic lupus erythematosus (SLE) is an autoimmune disorder marked by immune system dysregulation and autoantibodies production, causing widespread inflammation and damage across various body systems. Despite the prevalent back pain in SLE patients, the link between SLE and intervertebral disc (IVD) degeneration, a primary contributor to back pain, remains inadequately understood. This study explored the impact of SLE on IVD degeneration using the MRL/lpr mouse model, which effectively replicates human SLE manifestations.
Methods: The study utilized MRL/lpr mice to investigate the effects of SLE on IVD degeneration. The mice were evaluated for typical SLE phenotypes and indicators of IVD degeneration, including IVD height, IVD score, tissue integrity, extracellular matrix degradation, and apoptosis of IVD cells. Additionally, the study examined nucleus pulposus (NP) pyroptosis and inflammatory cytokine secretion. Mechanistic analysis focused on the antioxidant pathway, specifically the expression levels of NRF2, HO-1, KEAP1, and the phosphorylation levels of p65.
Results: MRL/lpr mice displayed typical SLE phenotypes and exacerbated profiles of IVD degeneration, including reduced IVD height, lower IVD score, significant IVD tissue impairment, extracellular matrix degradation, and increased apoptosis of IVD cells. Notably, SLE stimulated NP pyroptosis and excessive secretion of inflammatory cytokines. Mechanistic analysis indicated that the progression of SLE impedes the antioxidant pathway by downregulating NRF2 and HO-1 expression, upregulating KEAP1, and enhancing phosphorylation levels of p65.
Conclusion: Our findings highlight the mechanistic link between SLE and IVD degeneration, suggesting potential therapeutic targets for mitigating back pain in SLE patients.
Keywords: systemic lupus erythematosus, intervertebral disc degeneration, nucleus pulposus, pyroptosis, NRF2/KEAP1/NF-κB pathway
Introduction
Systemic lupus erythematosus (SLE) is a multifaceted autoimmune disease wherein immune system dysregulation mistakenly attacks healthy tissues and organs, predominantly affecting women of childbearing age.1 Manifesting through autoantibody/antigen complexes in diverse organs, SLE gives rise to a spectrum of systemic symptoms, encompassing conditions like lupus nephritis, lupus splenitis, arthritis, pericarditis, skin rashes, and organ inflammation.2–5 Apart from these typical symptoms, clinical evidence shows up to 16.3~36% of individuals with SLE encounter incidents of low back pain (LBP), adversely affecting their quality of life and daily performance.6–8 Despite extensive focus on the molecular intricacies of SLE pathogenesis and therapeutic modalities, the connection between SLE and LBP, predominantly caused by intervertebral disc (IVD) degeneration, remains incompletely understood.
IVD degeneration, a primary contributor to LBP, is characterized by metabolic disturbances within extracellular matrix (ECM), disc height reduction, structural impairment, inflammation, and decline in nucleus pulposus (NP) cell count.9,10 Notably, the synthesis of inflammatory cytokines such as IL-1β IL-18, IL-6, and TNF-α, have been identified as a key driver of IVD degeneration and consequent LBP.11 These cytokines amplify inflammatory cascades, accelerate the synthesis of other pro-inflammatory mediators such as cyclooxygenase-2 (COX-2), stimulate ECM degradation, hinder IVD cell proliferation, and increase oxidative stress throughout the degenerative process.12 Our recent findings from clinical evaluations of SLE-affected individuals and experimental studies using SLE model mice underscore heightened inflammatory cytokines within various tissues, including the cardiovascular system, and kidneys.13–16 Collectively, these studies suggest a correlation between excessive inflammation response within the context of SLE disorder and the progression of IVD degeneration.
Pyroptosis, an orchestrated form of programmed cell death, triggered via nod-like receptor protein-3 (NLRP3)/Caspase-1 activation, subsequently cleaving IL-1β and IL-18 precursors to spur their maturation and externalization, driving hyper-inflammatory reactions.17 Clinical and basic studies highlight the aberrant activation of pyroptosis in kidney tissue of lupus nephritis patients and SLE model mice,18,19 as well as within bone marrow-derived mesenchymal stem cells and monocytes/macrophages sourced from SLE patients.20,21 Meanwhile, anti-dsDNA antibodies, hallmarks of SLE, can activate the NLRP3 inflammasome in monocytes/macrophages of SLE patients, upregulating the production of IL-1β.22 Notably, inhibiting pyroptosis has shown effectiveness in alleviating lupus nephritis progression in both in vitro and in vivo settings.18,23 Moreover, SLE-induced pulmonary arterial hypertension has been linked to pyroptosis of pulmonary arterial endothelial cells.24 Given the role of NP cell pyroptosis in IVD degeneration and subsequent LBP.12,25 It is plausible to hypothesize that NP cell pyroptosis may also be involved in SLE-induced IVD degeneration.
Oxidative stress is a significant factor in the pathogenesis of SLE and other autoimmune diseases.26 The NRF2/KEAP1 pathway stands as a critical regulator of cellular antioxidant status. Upon oxidative stress, Nuclear factor erythroid 2-related factor 2 (NRF2) dissociates from Kelch-like ECH-associated protein 1 (KEAP1), translocates into the nucleus, and promotes the transcription of antioxidant genes such as heme oxygenase 1 (HO-1), thereby maintaining cellular homeostasis and preventing oxidative damage.27–29 Numerous studies on SLE patients and lupus-prone mice highlight the critical involvement of the NRF2/KEAP1 pathway in the SLE’s onset and progression and its associated phenotypes, such as glomerulonephritis.26,30–33 Moreover, mounting evidence shows the regulatory role of NRF2/KEAP1 pathway and downstream NF-κB signaling in the modulation of NP cell functions, including its pyroptosis activity, thereby governing the progression of IVD degeneration.34,35 Nonetheless, it remains unclear whether SLE induces NP cell pyroptosis to aggravate IVD degeneration by suppressing the NRF2/KEAP1 antioxidant pathway.
Given the significant burden of LBP on SLE patients, we aimed to explore the underlying mechanism of IVD degeneration coinciding with SLE progression. MRL/lpr lupus-prone mice, along their MRL/MpJ control compartments, were utilized, and a repertoire of investigative measures were employed, including radiography, morphological staining, immunohistological analyses, and TUNEL assay, to assess the complex interplay between SLE and IVD degeneration. Our discoveries provide an innovative understanding surrounding SLE-centric IVD degeneration and LBP intricacies.
Materials and Methods
Animals and Treatments
The MRL/lpr mouse model is renowned for its spontaneous lupus-like symptoms, including autoantibodies production (IgG anti-dsDNA and antiphospholipid antibodies), elevated cytokines, and clinical features such as hematological abnormalities, splenomegaly, and immune complex glomerulonephritis. In contrast, the genetically similar MRL/MpJ strain also eventually develops autoimmune disease but exhibits a healthy phenotype at 6 weeks of age, fully manifesting autoimmune traits by 24 weeks.13,15 In this study, female MRL/lpr mice and MRL/MpJ mice at 6 weeks of age, were obtained from the Center Animal House of Zhejiang Chinese Medical University and housed under a specific pathogen-free environment, and maintained at 23 ± 2°C, 40–60% humidity, and a 12-h light/dark cycle, with unrestricted access to food and water. All procedures were approved by the Ethics Committee for the Use of Experimental Animals at Zhejiang Chinese Medical University (No. IACUC-20211101-04).
At 6 weeks of age, female MRL/lpr mice were designated as the MRL/lpr group, and age-matched female MRL/MpJ mice served as controls. At 14 weeks of age, urine samples were collected using metabolic cages. Following collection, all mice were euthanized with 0.3% sodium pentobarbital. Blood was collected via abdominal aortic puncture, and the serum was separated and stored at −80°C until further measurement. Then, spleens, bilateral axillary and inguinal lymph nodes, right kidneys, and lumbar vertebrae were harvested for subsequent histopathological analysis. All animal experiments were obedient to the ARRIVE guidelines and conducted in accordance with the UK Animals (Scientific Procedures) Act, 1986 and related guidelines, EU Directive 2010/63/EU for animal experiments.36
Measurement of Spleen Index and Lymph Node Index
Spleen index was determined by the ratio of spleen weight to body weight, expressed as a percentage (Spleen Weight / Body Weight × 100%). Similarly, the lymph node index was calculated as the ratio of lymph node weight to body weight, also expressed as a percentage (Lymph Node Weight / Body Weight × 100%).
Analysis of Serum Indicators
The alterations of anti-ds-DNA, IL-10, and TNF-α levels in the serum of mice were measured using corresponding ELISA kits (Hnybio, Shanghai, China), according to the manufacturer’s instructions.
Analysis of Urine Protein
All fresh urine samples were centrifuged at 600 g for 15 min at 4 °C. The protein concentration in the supernatant was assessed using an automatic biochemical analyzer (TOSHIBA TBA-120FR).
Histology, Immunohistochemical (IHC) and Immunofluorescent (IF) Analyses
The renal histopathological evaluation was conducted according to the previous methods,13,15 kidney tissues were fixed in 4% buffered paraformaldehyde for 24 h and embedded in paraffin. Similarly, lumbar tissues were fixed in 4% buffered paraformaldehyde for 72 h, decalcified in 0.5 M ethylenediaminetetraacetic acid (EDTA; pH = 7.4) at room temperature for 3 weeks, dehydrated, and then embedded in paraffin. The kidney and lumbar tissues were sectioned into 5-mm-thick coronal slices, and the histology of these tissues was analyzed using hematoxylin-eosin (H&E) staining as previously described.13,37
Renal sections were incubated with primary antibodies to IgG (1:200, Abcam, Cambridge, UK) and C3 (1:200, Abcam). IVD sections were incubated with COL2 (1:300, NeoMarkers), AGGRECAN (1:300, Abcam), ADAMTS-5 (1:300, Abcam), MMP3 (1:300, Ruiying biological, Suzhou, China), BCL2 (1:300, Ruiying biological, Suzhou, China), BAX (1:300, Ruiying biological), CASPASE-3 (1:300, Ruiying biological), IL-1β (1:300, Bioss, Woburn, MA, USA), IL-18 (1:300, Ruiying biological), IL-6 (1:300, Proteintech, Wuhan, China), TNF-α (1:300, Ruiying biological), COX-2 (1:300, Proteintech), NLRP3 (1:300, Proteintech), ASC (1:300, Bioss), CASPASE-1 (1:300, Proteintech), NRF2 (1:300, ImmunoWay, Plano, TX, USA), KEAP1 (1:300, ImmunoWay), HO-1 (1:300, Proteintech), p-p65 (1:300, Cell Signaling, Danvers, MA, USA), p65 (1:300, Cell Signaling) at 4°C overnight. For IHC staining, sections were incubated with polymer-HRP labeled secondary antibody (ZSGB-BIO, Beijing, China) for 30 min the following day, and visualized using 0.05% diaminobenzidine (DAB). Positive staining appeared in brown, followed by counterstaining with hematoxylin. Images were captured with a microscope (Carl Zeiss, Göttingen, Germany). For IF analysis, a fluorescent-conjugated secondary antibody (Sungene Biotech, Tianjin, China) was added for 30 min in the dark, followed by DAPI counterstaining. The images were captured with a fluorescence microscope (Carl Zeiss, Göttingen, Germany). Quantitative histomorphometric analysis was conducted in a blinded manner with Image-Pro Plus Software version 6.0 (Media Cybernetics Inc, Rockville, Maryland, USA).
Micro-CT (μCT) Analysis
Before histological processing, the radiographic alterations in vertebrae were evaluated using high-resolution μCT analysis. A Skyscan 1176 μCT scanner (Skyscan 1176; Bruker μCT, Kontich, Belgium) with a 90 kVp source and 300 μA current was used to capture images of the vertebrae at a resolution of 9 μm. CTVol v2.2 visualization software was utilized to analyze various parameters of the L4-L5 intervertebral disc (IVD) and the intervertebral disc height index (DHI) was determined by measuring the anterior, middle, and posterior distances of the L4~L5 IVD and comparing them to the average height of the adjacent upper and lower vertebral body.
TUNEL Assay
The presence of apoptotic cells in situ was assessed using the TUNEL Bright Green Apoptosis Detection Kit (Vazyme Biotech; Nanjing, China), following the manufacturer’s instructions. To ensure specificity, PBS was used instead of TdT enzyme solution as a negative control. Six samples were randomly selected from each group, and the number of positive cells was quantified in each. Nuclei were identified separately by DAPI staining, and the total number of cells in each sample was counted to determine the percentage of cells that were undergoing apoptosis.
Cell Culture
Rat NP cell line (rNPCs) was generously gifted by Prof. Di Chen at Rush University Medical Center. The rNPCs were cultured in DMEM (Invitrogen) supplemented with 15% FBS (Ausbian, Australia), 2 mm L-glutamine (Invitrogen), and 0.1 mm non-essential amino acids (Invitrogen) in a humidified atmosphere of 5% CO2 at 37°C. The medium was renewed every 2 days. rNPCs were treated with LPS (5 μg/mL, L2880, Sigma-Aldrich), with or without TBHQ (5 μm, HY100489, MedChemExpress) for 12 h.
Western Blot Analysis
RIPA lysis buffer (Beyotime, Shanghai, China) containing 1% protease inhibitors was employed to extract the total protein of rNPCs. After 30 min-extraction at 4°C, the protein concentration was assessed using a Pierce™ BCA Protein assay kit (Thermo Fisher Scientific, Waltham, MA, USA). Subsequently, 30 μg protein was separated via SDS-PAGE and transferred onto nitrocellulose filter membranes (Millipore, Billerica, USA). After blocking with 5% non-fat milk for 1 h, the membranes were incubated with primary antibodies against NRF2 (Ser40) (p-NRF2) (1:1000, Bioss), NLRP3 (1:1000, Proteintech), IL-1β (1:1000, Bioss), IL-18 (1:1000, Bioss), COL2 (1:1000, Ruiying), MMP3 (1:1000, Bioss), and GAPDH (1:1000, Origen), at 4°C overnight, respectively. Subsequently, IRDye 680 or IRDye 800 secondary antibody was added and incubated for 2 hours. Signal visualization was achieved using the Odyssey Infrared Imaging System. GAPDH was used as the internal standard for total target proteins, and the blot intensities were quantified using Image J V1.8 (N ational Institutes of Health, Bethesda, MD, USA).
Statistical Analysis
All numerical data were presented as means ± SEM. Statistical analysis was performed using GraphPad Prism software (San Diego, CA, United States). Statistical comparisons between two groups were performed using the unpaired two-tailed Student’s t-test after confirming homogeneity of variances with the F-test with a p-value of less than 0.05 considered statistically significant.
Results
Verification of SLE Phenotypes in MRL/Lpr Mice
To gain a comprehensive understanding of the intricate relationship between SLE and IVD degeneration, MRL/MpJ and MRL/lpr mice were bred according to our established protocol.13 The onset and/or outcome of SLE in MRL/MpJ mice was initially confirmed by evaluating the ratio of spleen-to-mouse body weight and the ratio of lymph node-to-mouse body weight, two representative indexes of inflammatory response that reflect the severity of disease to some extent. The results revealed increased ratios of spleen index and lymph node index in MRL/lpr mice compared to MRL/MpJ mice (Figure 1A and B). In parallel, MRL/lpr mice exhibited heightened serum levels of dsDNA, IL-10, and TNF-α (Figure 1C–E), as well as elevated urinary protein levels (Figure 1F). Furthermore, Histopathological analysis using H&E staining revealed substantial kidney impairments in the MRL/lpr mice, including mesangial matrix hyperplasia, mesangial cell proliferation, glomerular swelling, and inflammatory cell infiltration (Figure 1G). Further IF analysis of IgG and IHC analysis of C3 indicated increased IgG and C3 deposition in their glomeruli (Figure 1H–K). All these data demonstrate the onset and exacerbation of SLE within the MRL/lpr mouse at 14 weeks of age.
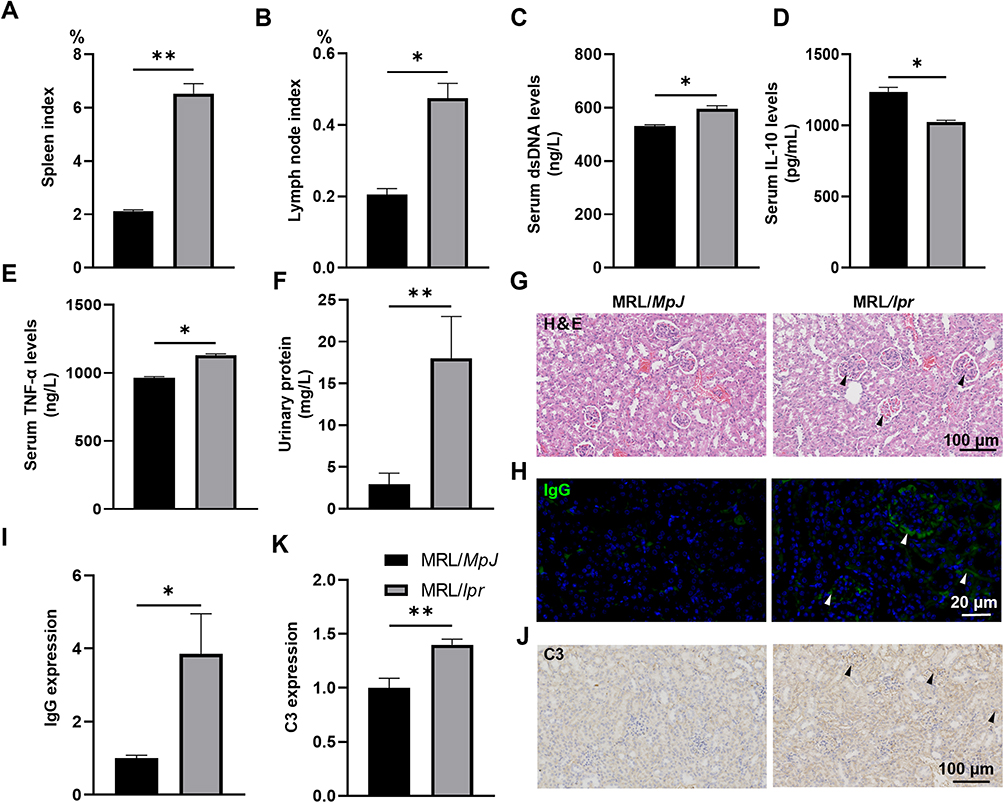 |
Figure 1 Verification of SLE phenotypes in MRL/lpr mice. (A) Spleen indexes in MRL/lpr mice and MRL/MpJ mice were calculated as the ratio of spleen weight to body weight (%). (B) Lymph node indexes were calculated as the ratio of lymph node weight to body weight (%). (C–E) Serum levels of anti-dsDNA antibody (C), IL-10 (D), and TNF-α (E) in MRL/lpr mice and MRL/MpJ mice were determined by corresponding ELISA kits. (F) Quantification of urinary protein concentration in MRL/lpr mice and MRL/MpJ mice. (G) Representative images of the kidney H&E staining sections. (H and I) Immunofluorescence staining (H) and corresponding quantification (I) of IgG deposition in the glomeruli of MRL/lpr mice. (J and K) IHC analysis (J) and corresponding quantification (K) of C3 deposition in the glomeruli of MRL/lpr mice. Data are presented as mean ± SEM. *p < 0.05; **p < 0.01 (vs MRL/MpJ mice), n = 6 per group. Abbreviation: SLE, Systemic lupus erythematosus. |
SLE Contributes to IVD Degeneration of MRL/Lpr Mice
To investigate the impact of SLE on the development of IVD degeneration, a comparative analysis of radiographic changes in L4-L5 IVDs of MRL/MpJ mice and MRL/lpr mice was conducted using μCT analysis. The DHI results showed that MRL/lpr mice displayed a 20% reduction in lumbar disc height compared to MRL/MpJ mice (Figure 2A). To determine whether SLE could affect the structure and composition of IVDs in MRL/lpr mice, histopathological assessment of IVDs was carried out using H&E staining and a histological score system based on specific histological grading scale criteria as previously described.38 The corresponding results showed that MRL/MpJ mice displayed well-organized IVD structures, characterized by ample vacuoles with abundant matrix contents in NP, orderly arranged annulus fibrosus (AF) tissues with minimal tearing, and no ectopic bone formation in endplates (EP). Strikingly, the IVDs of MRL/lpr mice exhibited reduced vacuole sizes in NP, fissures along with folds in the interlamellar regions of AF tissues, and extensive ectopic bone formation in EP (Figure 2B). In parallel, the histological grading system revealed notably lower histological scores for IVDs of MRL/lpr mice (Figure 2C). Our findings suggest that SLE contributes to the deterioration of IVD structure in MRL/lpr mice.
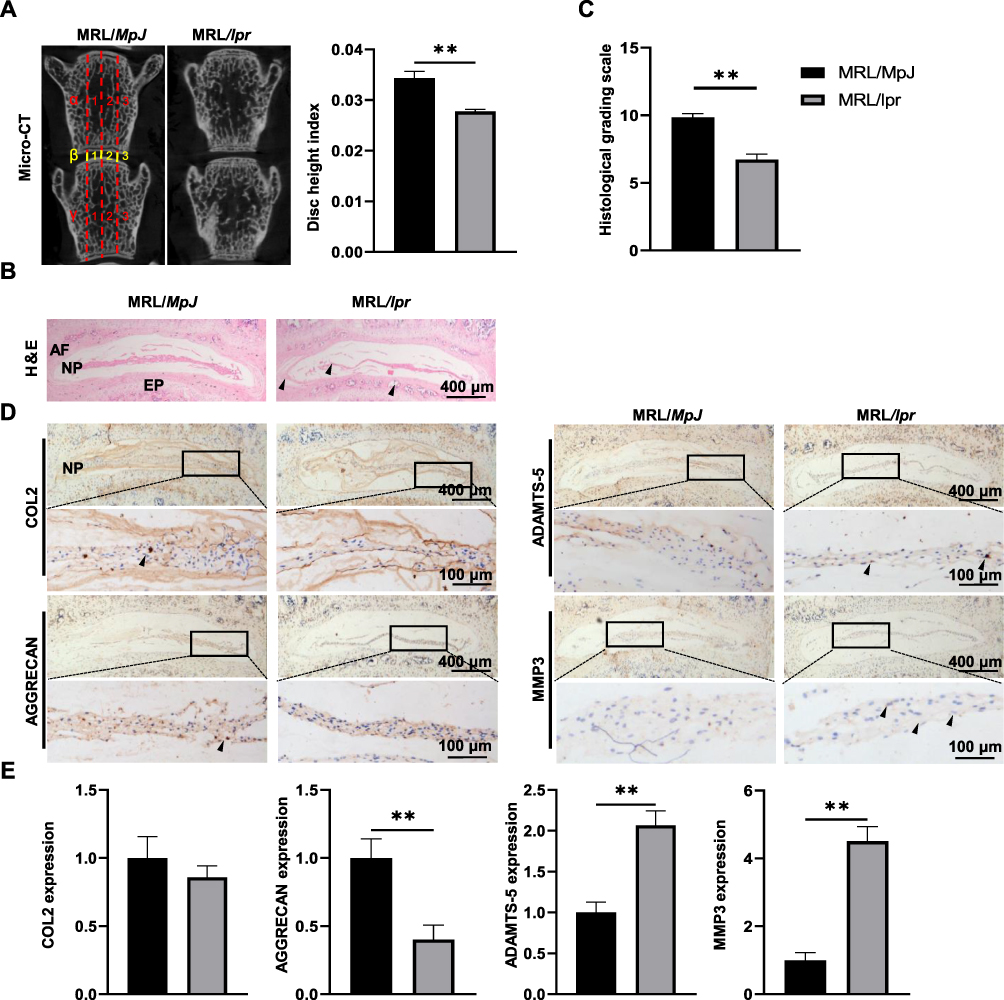 |
Figure 2 SLE contributes to IVD degeneration in MRL/lpr mice. (A) Representative μCT images of IVDs between L4 and L5 vertebrae of MRL/MpJ and MRL/lpr mice. The DHI was calculated based on measurements of adjacent L4 and L5 vertebrae. Yellow lines represent the distances (β1-β3) between the adjacent vertebra, and red lines represent adjacent vertebral body heights (α1-α3 and γ1-γ3). (B) Representative images of the H&E staining results of IVD tissues in MRL/MpJ and MRL/lpr mice. (C) Evaluation of IVD degeneration degree in (B) by histological grading score. (D and E) The IHC staining (D) and corresponding quantification (E) of COL2, AGGRECAN, ADAMTS-5, and MMP3 in IVDs. Black triangles indicate positive staining cells. Data are expressed as mean ± SEM, **p < 0.01 (vs MRL/MpJ mice), n = 6 per group. Abbreviations: SLE, Systemic lupus erythematosus; μCT, Micro-CT; L4, Lumbar 4; IHC, Immunohistochemical analysis. |
SLE Disrupts the Homeostasis of Matrix Metabolism in IVDs of MRL/Lpr Mice
It has been well established that maintaining a stable matrix composition is paramount for the proper physiological functioning of IVDs.39 To assess the effect of SLE on ECM metabolism of IVDs, the expression levels of AGGRECAN and COL2, and their corresponding degradases ADAMTS-5 and MMP3, were determined using IHC analysis. As depicted in Figure 2D, MRL/lpr mice exhibited a 60% reduction in AGGRECAN levels and a 2.1-fold and 4.5-fold increase in ADAMTS5 and MMP3 within the IVD, albeit with a non-significant decrease in COL2. These findings reveal that SLE contributes to ECM degradation within IVDs, thereby aggravating IVD degeneration.
SLE Augments Apoptosis Ratio in NP Tissues of MRL/lpr Mice
To identify the impact of SLE on the apoptosis of IVD cells, the expression of apoptotic-related markers, BCL2, BAX, and CASPASE-3 were determined using IF analysis. We found a significant down-regulation of anti-apoptotic protein BCL2 and up-regulation of pro-apoptotic proteins BAX and CASPASE-3 in IVDs of MRL/lpr mice (Figure 3A–D). Similarly, the pro-apoptosis effect of SLE was further confirmed by TUNEL staining, showing a notable rise in the number of TUNEL-positive cells within NP (Figure 3E and F). The above results suggest that the exacerbation of SLE could potentiate apoptosis in IVD tissues of MRL/lpr mice.
 |
Figure 3 Exacerbation of SLE enhances apoptosis rate in IVDs of MRL/lpr mice. (A–D) IF staining (A) and corresponding quantification (B–D) for the expression of BCL2, BAX, and CASPASE-3 in IVDs of MRL/MpJ and MRL/lpr mice. White triangles indicate the high expression in A. DAPI stains nuclei blue. (E and F) TUNEL staining (E) and corresponding quantification of TUNEL-positive cell rates (F) in the IVD tissues. White triangles indicate TUNEL-positive cells in (E). DAPI stains nuclei blue. Data are expressed as mean ± SEM, **p < 0.01 (vs MRL/MpJ mice), n = 6 per group. Abbreviations: SLE, Systemic lupus erythematosus; IVD, Intervertebral disc; IF, Immunofluorescent analysis. |
Exacerbation of SLE Facilitates Inflammatory Response in NP Tissues of MRL/Lpr Mice
To explore whether SLE affects the inflammatory response within IVDs of MRL/lpr mice, the expressions of inflammatory cytokines, including IL-1β, IL-18, IL-6, TNF-α, and COX2, were determined by IF assay. Remarkably, the progression of SLE amplified the production of these inflammatory cytokines in IVDs of MRL/lpr mice (Figure 4). Specifically, the expression levels of IL-1β, IL-18, IL-6, TNF-α, and COX2 were found to be 1.9-fold, 10.1-fold, 2.8-fold, 4.1-fold, and 1.8-fold higher, respectively, in the NP tissues of MRL/lpr mice compared to MRL/MpJ mice, suggesting a robust inflammatory response within IVDs following the exacerbation of SLE.
 |
Figure 4 Exacerbation of SLE promotes inflammatory response in NP tissues of MRL/lpr mice. (A) IF staining for the expression of IL-1β, IL-18, IL-6, TNF-α, and COX-2 in NP tissues of MRL/lpr mice compared to MRL/MpJ mice. White triangles indicate the high expression in NP. DAPI stains nuclei blue. (B) Quantification of the expression of IL-1β, IL-18, IL-6, TNF-α, and COX-2 in. (A) Data are expressed as mean ± SEM, **p < 0.01 (vs MRL/MpJ mice), n = 6 per group. Abbreviations: SLE, Systemic lupus erythematosus; NP, Nucleus pulposus; IF, Immunofluorescent analysis; COX-2, Cyclooxygenase-2. |
Exacerbation of SLE Aggravates Pyroptosis of NP Tissues of MRL/Lpr Mice
Considering the well-established involvement of IL-1β and IL-18 in the pathology of IVD degeneration and its maturation through inflammasomes, along with the widely recognized role of the molecular mechanism underlying NLRP3 inflammasome-mediated NP pyroptosis during IVD degeneration progression,12,25,37,40 we further evaluated the impact of SLE on the pyroptosis activity in IVDs by determining the key pyroptosis-related proteins including NLRP3, ASC, and CASPASE-1 using IF analyses. As expected, there was a 2.4-fold, 2.2-fold, and 5.7-fold upregulation in the expression levels of NLRP3, ASC, and CASPASE-1, respectively, in the NP tissues of MRL/lpr mice (Figure 5). The findings indicate that the advancement of SLE aggravates inflammation response primarily through stimulating NLRP3-mediated NP pyroptosis.
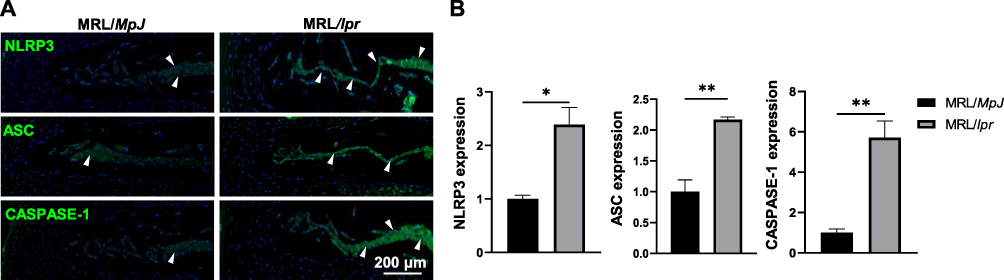 |
Figure 5 SLE activates NLRP3-mediated pyroptosis in NP tissues of MRL/lpr mice. (A) IF staining for the expression of NLRP3, ASC, and CASPASE-1 in NP tissues of MRL/lpr mice compared to MRL/MpJ mice. White triangles indicate the high expression in NP. DAPI stains nuclei blue. (B) Quantification of the expression of NLRP3, ASC, and CASPASE-1 in (A). Data are expressed as mean ± SEM, *p < 0.05, **p < 0.01 (vs MRL/MpJ mice), n = 6 per group. Abbreviations: SLE, Systemic lupus erythematosus; NLRP3, Nod-like receptor protein-3; NP, Nucleus pulposus; IF, Immunofluorescent analysis. |
Exacerbation of SLE Suppresses NRF2/KEAP1/NF-κB Pathway in NP Tissues of MRL/Lpr Mice
Subsequently, to elucidate the underlying mechanism through which SLE activates NP pyroptosis, we assessed the NRF2/KEAP1/NF-κB signaling pathway by analyzing the expression levels of NRF2, KEAP1, HO-1, as well as the phosphorylation status of p65—key upstream regulators of NP pyroptosis, using IF analyses. As illustrated in Figure 6, SLE significantly reduced the expression of NRF2 and HO-1 expressions, while increasing the levels of KEAP1, p-p65 and total p65 expressions. To further validate the crucial role of the NRF2-KEAP1 pathway in NP pyroptosis in vitro, we treated rNPCs with a NRF2 agonist tert-Butylhydroquinone (TBHQ) in the presence of LPS, a classic pyroptosis inducer.41 As expected, TBHQ treatment significantly reversed LPS-induced decrease in NRF2 phosphorylation and the increase in NLRP3 and its downstream cytokines, IL-1β and IL-18, in NP cells, along with the restoration of matrix metabolism. Given the well-established correlation between the oxidative stress status in SLE,30 and the interplay between oxidative stress, NRF2/KEAP1/NF-κB signaling pathway, and NP pyroptosis, these findings collectively suggest that SLE may potentiate NP pyroptosis via governing NRF2/KEAP1/NF-κB signaling pathway, thereby exacerbating IVD degeneration progression (Figure 7).
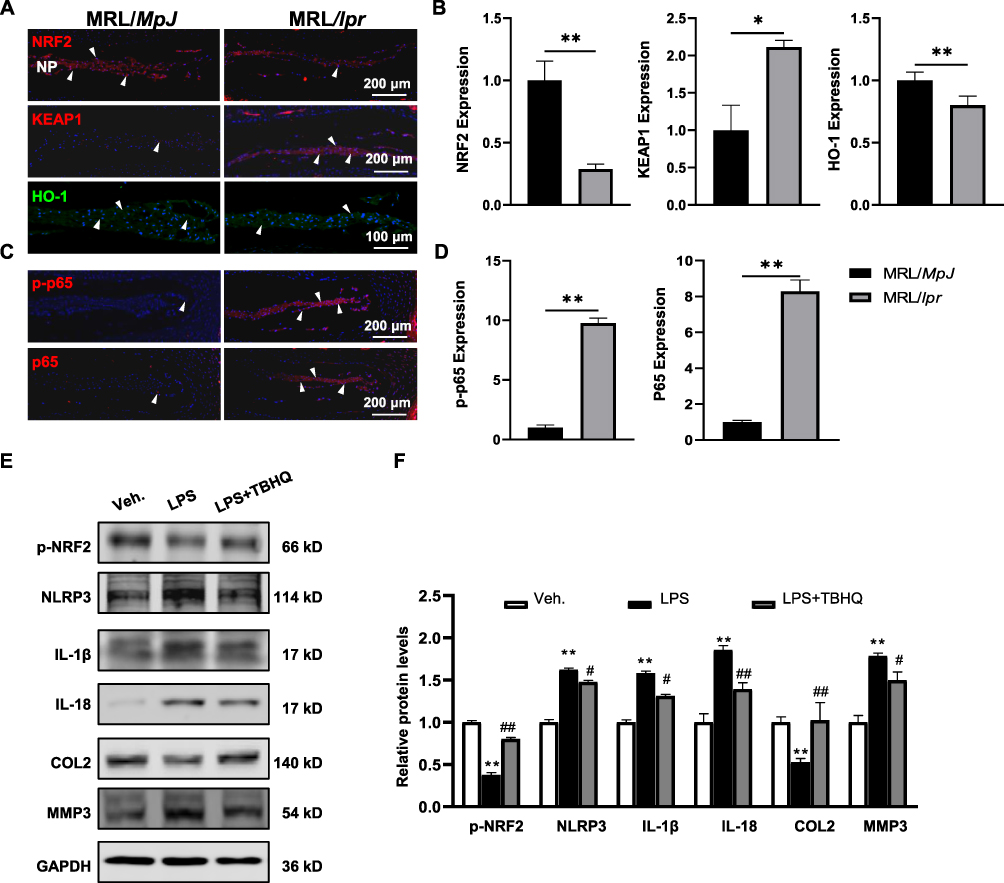 |
Figure 6 Exacerbation of SLE regulates NRF2/KEAP1/NF-κB pathway in NP tissues of MRL/lpr mice. (A) IF staining for the expression of NRF2, KEAP1, and HO-1, and corresponding quantification (B) in the NP tissues of MRL/lpr mice compared to MRL/MpJ mice. (C) IF staining for the expression of p-p65 and p65, and corresponding quantification (D) in the NP of mice. White triangles indicate the high expression in NP. DAPI stains nuclei blue. Data are expressed as mean ± SEM, *p < 0.05, **p < 0.01 (vs MRL/MpJ mice), n = 6 per group. (E) Western blot analysis of p-NRF2, NLPR3, IL-1β, IL-18, COL2, and MMP3 in rNPCs treated with LPS and TBHQ for 12 hours. (F) Quantification of relative protein levels in (E). Data are expressed as mean ± SEM, **p < 0.01 (vs rNPCs treated with Vehicle (Veh.)), #p < 0.05, ##p < 0.01, (vs rNPCs treated with LPS alone). Abbreviations: SLE, Systemic lupus erythematosus; NRF2, Nuclear factor E2-related factor 2; KEAP1, Kelch-like ECH-associated protein 1; HO-1, Heme oxygenase 1; p-p65, phosphorylated p65. |
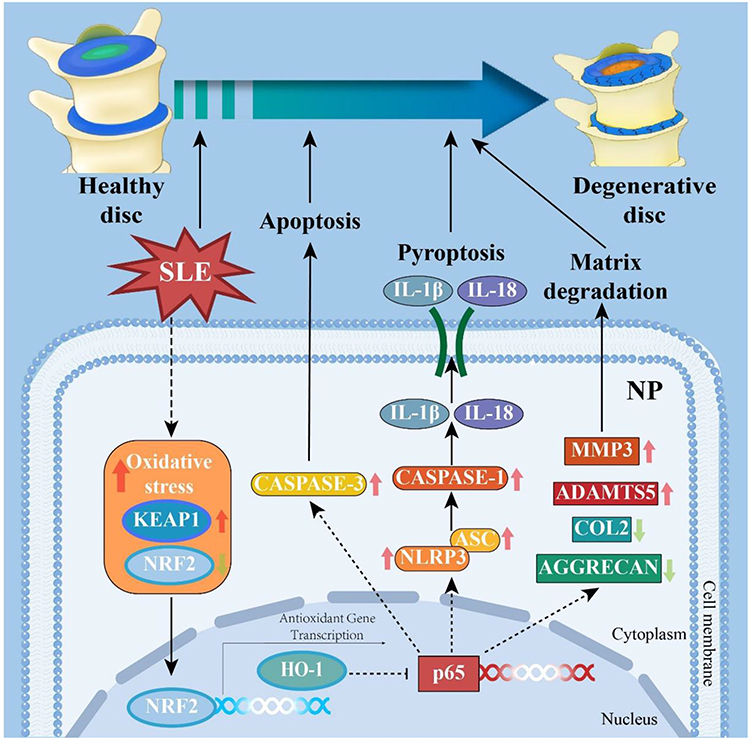 |
Figure 7 Schematic working model of SLE strengthens NP cell pyroptosis and IVD degeneration via regulation of NRF2/KEAP1/NF-κB pathway. Abbreviations: SLE, Systemic lupus erythematosus; NP, Nucleus pulposus; IVD, Intervertebral disc; NRF2, Nuclear factor E2-related factor 2; KEAP1, Kelch-like ECH-associated protein 1. |
Discussion
SLE is a complex autoimmune disease with multisystem involvement, characterized by the formation of autoantibodies, deposition of immune complexes, and inflammation, leading to a spectrum of complications, presents notably with cutaneous and mucosal complications (70%), renal complications (30%), and especially arthritis and musculoskeletal complications (85%).42 Notably, accumulating studies on SLE populations have underscored that approximately 16.3~36% of SLE patients encounter persistent LBP,6,8 indicating a heightened incidence of IVD degeneration in the context of SLE disorder. Despite this, the precise mechanism linking IVD degeneration advancement to SLE exacerbation remains obscure. In the present study, a comprehensive comparison analysis was conducted utilizing 14-week-old MRL/MpJ and MRL/lpr mice to assess the potential impact and the underlying mechanism of SLE occurrence on IVD degeneration pathogenesis. Our findings revealed that MRL/lpr mice displayed typical SLE manifestations, alongside compromised IVD homeostasis including reduced IVD height, diminished vacuole sizes in NP, ectopic bone formation in EP, accelerated ECM degradation, increased apoptosis ratio of IVD cells, and excessive secretion of inflammatory cytokines. Importantly, further analysis indicated that SLE stimulated NP pyroptosis by upregulation of the key pyroptosis-related proteins, including NLRP3, ASC, and CASPASE-1, via suppressing antioxidant pathway, NRF2/KEAP1 pathway and enhancing NF-κB signaling. Overall, our in vivo findings enhance understanding concerning how SLE stimulates LBP by promoting NP pyroptosis to exacerbate IVD degeneration development, highlighting a promising therapeutic target for improving the clinical management of LBP in SLE patients.
MRL/lpr lupus-prone mice, with accelerated autoimmune responses due to Fas (CD95) mutations, mirroring key pathological features of human autoimmune diseases and related manifestations, including lupus nephritis (glomerulonephritis, immune complex deposition, proteinuria), autoantibody production (ANA, anti-dsDNA, and anti-Sm), cutaneous lesions (ulcers, alopecia, dermatitis), lymphadenopathy, splenomegaly, arthritis (polyarthritis), joint inflammation and cartilage degeneration, and hematological abnormalities (hemolytic anemia, thrombocytopenia),43–45 making them as invaluable assets for investigating the complex interplay of inflammatory responses and oxidative stress in various systemic manifestations associated with SLE.23 Concurrently, recent evidence highlights that approximately 90% of SLE patients experienced joint symptoms ranging from intermittent arthralgias to acute polyarthritis, with a significant portion suffering from LBP.8 Consistent with these findings, in the present study, our investigation on IVD homeostasis using MRL/lpr mice showed that the onset of SLE triggers the development of IVD degeneration, as evidenced by reduced IVD height, impaired IVD structure, accelerated ECM degradation, activated inflammatory response, and loss of NP cells within IVD tissues, solidifying that SLE is responsible for the progression of IVD degeneration and subsequent LBP.
Numerous studies have confirmed that SLE induces congregation of many lupus autoantigens inside apoptotic cells and excessive production of inflammatory cytokines in various tissues, which effectively regulate the phenotypes of SLE manifestations such as renal injury,46 and cardiovascular system.47,48 It is worth noting that the increased serum IL-18 levels in SLE patients were closely associated with the severity of lupus nephritis,49 while our latest findings using MRL/lpr mice suggest that excessive inflammation in cardiac tissue and renal tissue triggers the aberrant lipid metabolism, thereby facilitating the progression of lupus nephritis and premature cardiovascular disease.13,14 Herein, our findings provide evidence that SLE leads to enhanced productions of inflammatory factors in IVD tissues, notably a 10.1-fold elevation of IL-18, in turn, contributes to ECM degradation, and apoptosis of IVD cells, thus promoting the occurrence of IVD degeneration. Importantly, while apoptosis typically does not elicit an inflammatory response in organisms, pyroptosis acts more rapidly than apoptosis and is associated with a substantial release of pro-inflammatory factors.50 Therefore, we speculate that SLE-induced IVD degeneration may not be solely attributed to apoptosis but rather to the excessive production of cytokines induced by pyroptosis.
Pyroptosis plays a crucial role in initiating the inflammatory response, leading to the significant accumulation and release of cytokines IL-1β and IL-18.51 Several studies have reported that polymorphisms in pyroptosis-related gene are associated with the susceptibility, disease severity, and treatment effect of SLE.52–54 For instance, Cruz et al explored the distribution of 13 SNPs in eight pyroptosis-related genes and found that the NLRP3 gain-of-function variant (rs10754558) was significantly more frequent in SLE patients with nephritis,54 while Chen et al also discovered that the IL-18 gene promoter rs1946518 (−607A/C) polymorphism was closely associated with susceptibility to SLE.53 Moreover, a multitude of investigations both in SLE individuals and mice have demonstrated that aberrant activation of pyroptosis was noted in glomerular mesangial cells, podocytes, macrophage, and CD4+ T cells,18,23,54–59 while inhibiting pyroptosis has thus emerged as a promising therapeutic strategy with the potential to significantly retard SLE progression.60–62 Consistent with the aforementioned activation of pyroptosis in SLE-related manifestations, our study unveils that SLE significantly triggers NP pyroptosis within IVD tissues of MRL/lpr mice, evidenced by the augmented expression of pyroptosis-related proteins including NLRP3, ASC, CASPASE-1. These findings reinforce the assertion that pyroptosis may play a crucial role, surpassing apoptosis, during SLE-induced IVD degeneration and LBP. Recent findings show that the DDRGK1-NRF2-KEAP1 interaction regulates redox balance and chemoresistance, offering new therapeutic prospects for osteosarcoma treatment.
The NRF2/KEAP1 pathway is the most important antioxidant system protecting cells from damage due to oxidative stress,63 and has been demonstrated to manage oxidative stress and inflammation in the progression of lupus-like symptoms and IVD degeneration.26,30–32,64 Upregulation of NRF2/HO-1 and suppression of NF-κB/NLRP3 pathway have shown beneficial effects in mitigating murine lupus nephritis,65 and IVD degeneration.66,67 Consistent with these findings, our results demonstrate the occurrence of SLE significantly decreases NRF2 and HO-1 while increasing KEAP1 expression, alongside upregulation of the phosphorylated and total levels of p65. These outcomes strongly suggest that SLE-mediated induction of NP cell pyroptosis may contribute to ECM degradation, apoptosis of IVD cells, inflammatory response, and impaired IVD structure via the regulation of the NRF2/KEAP1/NF-κB pathway.
While this study has yielded significant findings, several limitations must be acknowledged. This study exclusively utilized the MRL/lpr mouse model, which, although valuable, cannot represent the full spectrum of SLE pathology. Comparative studies using other lupus-prone mouse models, such as NZB/W F1,68 are necessary to ensure that the findings are broadly applicable and not limited to one specific model. This approach would validate the observed effects and enhance the generalizability of the results. Moreover, our study evaluated the mice at 14 weeks of age, focusing on a relatively short-term period. Given that both SLE and IVD degeneration are chronic conditions, it is crucial to understand the long-term effects and progression dynamics. Long-term studies examining older mice at various stages would provide a more comprehensive understanding of how SLE influences IVD degeneration over time, capturing the chronic and progressive nature of these diseases. Furthermore, despite the importance of animal models in understanding disease mechanisms, translating these findings to humans is crucial. Validation using clinical samples from SLE patients is necessary to verify if the molecular mechanisms observed in mice correspond to human pathology. This would strengthen the translational relevance of the findings and potentially guide clinical interventions. Addressing these limitations through future research will enhance our understanding and pave the way for more effective therapeutic strategies.
Conclusions
In conclusion, our work demonstrates that SLE exacerbates IVD degeneration by promoting NP cell pyroptosis and governing the NRF2/KEAP1/NF-κB pathway, offering new insights into SLE-associated LBP and suggesting potential targets for therapeutic intervention. Future research should validate these results in diverse models and human samples to enhance translational relevance.
Ethics Approval
The study was conducted in accordance with the Declaration of Helsinki, and all procedures were approved by the Ethics Committee for the Use of Experimental Animals at Zhejiang Chinese Medical University (Approval No. IACUC-20211101-04).
Acknowledgments
This research was funded by Natural Science Foundation of Zhejiang Province (No. LY22H270003, LQ23H270003), National Natural Science Foundation of China (No. 82174140, 82174401, 81973870), the Joint Funds of the Zhejiang Provincial Natural Science Foundation of China under Grant No. LBY22H270008 and BY21H060010, Traditional Chinese Medical Administration of Zhejiang Province (No. 2023ZR019, 2023ZL128, 2022ZX005, 2022ZB119, 2021ZB090), Zhejiang medical and health science and technology project (No. 2023RC194, 2021KY222), Research Project of Zhejiang Chinese Medical University Scientific (No. 2023JKZKTS27 and 2021JKZDZC02), Research Project of Zhejiang Chinese Medical University Affiliated Hospital (No. 2022FSYYZZ05 and 2022FSYYZQ02). We are grateful to Xiaoping Xu and Dejun Wang at the Center Animal House of Zhejiang Chinese Medical University for their expert care and management of the mice. Their dedication and invaluable assistance were crucial to the successful conduct of this study. We deeply appreciate their professionalism and commitment to ensuring the well-being of the animals, which significantly contributed to the reliability and success of our research.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Almaani S, Meara A, Rovin BH. Update on Lupus Nephritis. Clin J Am Soc Nephrol. 2017;12(5):825–835. doi:10.2215/CJN.05780616
2. Tsokos GC. Systemic lupus erythematosus. N Engl J Med. 2011;365(22):2110–2121. doi:10.1056/NEJMra1100359
3. Xu Q, Zhang X, Ge S, Xu C, Lv Y, Shuai Z. Triptoquinone A and B exercise a therapeutic effect in systemic lupus erythematosus by regulating NLRC3. PeerJ. 2023;11:e15395.
4. Fan Y, Hao Y-J, Zhang Z-L. Systemic lupus erythematosus: year in review 2019. Chin Med J. 2020;133(18):2189–2196. doi:10.1097/CM9.0000000000000983
5. Reiss AB, Jacob B, Ahmed S, Carsons SE, DeLeon J. Understanding accelerated atherosclerosis in systemic lupus erythematosus: toward better treatment and prevention. Inflammation. 2021;44(5):1663–1682. doi:10.1007/s10753-021-01455-6
6. Figueroa-Parra G, Jeffery MM, Dabit JY, et al. Long-term opioid therapy among patients with systemic lupus erythematosus in the community: a lupus midwest network (LUMEN) study. J Rheumatol. 2023;50(4):504–511. doi:10.3899/jrheum.220822
7. Gupta V, Dhanota DS. Symptomatic bilateral sacroiliitis in a patient with juvenile systemic lupus erythematosus: a rare association. Medit J Rheumatol. 2023;34(1):105. doi:10.31138/mjr.34.1.105
8. Yilmaz N, Yazici A, Türkmen BÖ, Karalok I, Yavuz Ş. Sacroiliitis in systemic lupus erythematosus revisited. Archiv Rheumatol. 2020;35(2):254. doi:10.46497/ArchRheumatol.2020.7514
9. Sun K, Jiang J, Wang Y, et al. The role of nerve fibers and their neurotransmitters in regulating intervertebral disc degeneration. Ageing Res Rev. 2022;81:101733. doi:10.1016/j.arr.2022.101733
10. Tamagawa S, Sakai D, Nojiri H, et al. SOD2 orchestrates redox homeostasis in intervertebral discs: a novel insight into oxidative stress-mediated degeneration and therapeutic potential. Redox Biol. 2024;71:103091. doi:10.1016/j.redox.2024.103091
11. Zehra U, Tryfonidou M, Iatridis JC, Illien-Jünger S, Mwale F, Samartzis D. Mechanisms and clinical implications of intervertebral disc calcification. Nat Rev Rheumatol. 2022;18(6):352–362. doi:10.1038/s41584-022-00783-7
12. Ge Y, Chen Y, Guo C, et al. Pyroptosis and intervertebral disc degeneration: mechanistic insights and therapeutic implications. J Inflamm Res. 2022;Volume 15:5857–5871. doi:10.2147/JIR.S382069
13. Zhang J, Lu L, Tian X, et al. Lipidomics revealed aberrant lipid metabolism caused by inflammation in cardiac tissue in the early stage of systemic lupus erythematosus in a murine model. Metabolites. 2022;12(5):415. doi:10.3390/metabo12050415
14. Hu C, Du Y, Xu X, et al. Lipidomics revealed aberrant metabolism of lipids including FAHFAs in renal tissue in the progression of lupus nephritis in a murine model. Metabolites. 2021;11(3):142. doi:10.3390/metabo11030142
15. Hu C, Zhang J, Hong S, et al. Oxidative stress-induced aberrant lipid metabolism is an important causal factor for dysfunction of immunocytes from patients with systemic lupus erythematosus. Free Radic Biol Med. 2021;163:210–219. doi:10.1016/j.freeradbiomed.2020.12.006
16. Hu C, Zhou J, Yang S, et al. Oxidative stress leads to reduction of plasmalogen serving as a novel biomarker for systemic lupus erythematosus. Free Radic Biol Med. 2016;101:475–481. doi:10.1016/j.freeradbiomed.2016.11.006
17. Gram AM, Booty LM, Bryant CE. Chopping GSDMD: caspase-8 has joined the team of pyroptosis-mediating caspases. EMBO J. 2019;38:10. doi:10.15252/embj.2019102065
18. Cao H, Liang J, Liu J, et al. Novel effects of combination therapy through inhibition of caspase-1/gasdermin D induced-pyroptosis in lupus nephritis. Front Immunol. 2021;12:720877. doi:10.3389/fimmu.2021.720877
19. Huang Y, Yang -D-D, Li X-Y, Fang D-L, Zhou W-J. ZBP1 is a significant pyroptosis regulator for systemic lupus erythematosus. Ann Translat Med. 2021;9(24):1773. doi:10.21037/atm-21-6193
20. Tan W, Gu Z, Leng J, et al. Let-7f-5p ameliorates inflammation by targeting NLRP3 in bone marrow-derived mesenchymal stem cells in patients with systemic lupus erythematosus. Biomed Pharmacother. 2019;118:109313. doi:10.1016/j.biopha.2019.109313
21. Inokuchi S, Mitoma H, Kawano S, et al. Activation of caspase-1 is mediated by stimulation of interferon genes and NLR family pyrin domain containing 3 in monocytes of active systemic lupus erythematosus. Clin Exp Rheumatol. 2022;40(3):522–531. doi:10.55563/clinexprheumatol/eakvlv
22. Zhang H, Fu R, Guo C, et al. Anti-dsDNA antibodies bind to TLR4 and activate NLRP3 inflammasome in lupus monocytes/macrophages. J Transl Med. 2016;14:1–12. doi:10.1186/s12967-016-0911-z
23. Chen L, Li F, J-H N, et al. Ursolic acid alleviates lupus nephritis by suppressing SUMO1-mediated stabilization of NLRP3. Phytomedicine. 2024;130:155556. doi:10.1016/j.phymed.2024.155556
24. Xing Y, Zhao J, Zhou M, et al. The LPS induced pyroptosis exacerbates BMPR2 signaling deficiency to potentiate SLE‐PAH. THE FASEB Journal. 2021;35(12):e22044. doi:10.1096/fj.202100851RR
25. Zhan K, Zhu K, Gu B, et al. MINK1 deficiency stimulates nucleus pulposus cell pyroptosis and exacerbates intervertebral disc degeneration. Int Immunopharmacol. 2024;134:112202. doi:10.1016/j.intimp.2024.112202
26. Gautam P, Kaur G, Tandon A, Sharma A, Bhatnagar A. Altered redox regulation by Nrf2-Keap1 system in dendritic cells of systemic lupus erythematosus patients. Lupus. 2020;29(12):1544–1555. doi:10.1177/0961203320950022
27. Ali I, Li C, Kuang M, et al. Nrf2 Activation and NF-Kb & caspase/bax signaling inhibition by sodium butyrate alleviates LPS-induced cell injury in bovine mammary epithelial cells. Mol Immunol. 2022;148:54–67. doi:10.1016/j.molimm.2022.05.121
28. Jin X, Wang K, Liu H, Hu F, Zhao F, Liu J. Protection of bovine mammary epithelial cells from hydrogen peroxide-induced oxidative cell damage by resveratrol. Oxid Med Cell Longev. 2016;2016:2572175. doi:10.1155/2016/2572175
29. Sun HJ, Ding S, Guan DX, Ma LQ. Nrf2/Keap1 pathway in countering arsenic-induced oxidative stress in mice after chronic exposure at environmentally-relevant concentrations. Chemosphere. 2022;303(Pt 3):135256. doi:10.1016/j.chemosphere.2022.135256
30. Ohl K, Tenbrock K. Oxidative stress in SLE T Cells, Is NRF2 really the target to treat? Front Immunol. 2021;12:633845. doi:10.3389/fimmu.2021.633845
31. Gautam P, Kaushal J, Sharma A, Bhatnagar A. Proportion of B cell subsets and Nrf2 mediated redox regulation in systemic lupus erythematosus patients. Immunobiology. 2022;227(2):152180. doi:10.1016/j.imbio.2022.152180
32. Tandon A, Anupam K, Kaushal J, Gautam P, Sharma A, Bhatnagar A. Altered oxidative stress markers in relation to T cells, NK cells & killer immunoglobulin receptors that are associated with disease activity in SLE patients. Lupus. 2020;29(14):1831–1844. doi:10.1177/0961203320959441
33. Córdova E, Velázquez-Cruz R, Centeno F, Baca V, Orozco L. The NRF2 gene variant,-653G/A, is associated with nephritis in childhood-onset systemic lupus erythematosus. Lupus. 2010;19(10):1237–1242. doi:10.1177/0961203310367917
34. Xu J, Xie G, Yang W, Wang W, Zuo Z, Wang W. Platelet-rich plasma attenuates intervertebral disc degeneration via delivering miR-141-3p-containing exosomes. Cell Cycle. 2021;20(15):1487–1499. doi:10.1080/15384101.2021.1949839
35. Lu G, Zhang C, Li K, et al. Sinomenine Ameliorates IL-1β-induced intervertebral disc degeneration in rats through suppressing inflammation and oxidative stress via Keap1/Nrf2/NF-κB signaling pathways. J Inflamm Res;2023. 4777–4791. doi:10.2147/JIR.S430423
36. Kilkenny C, Browne WJ, Cuthill IC, Emerson M, Altman DG. Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. J Pharmacol Pharmacotherapeutics. 2010;1(2):94–99. doi:10.4103/0976-500X.72351
37. Fu F, Bao R, Yao S, et al. Aberrant spinal mechanical loading stress triggers intervertebral disc degeneration by inducing pyroptosis and nerve ingrowth. Sci Rep. 2021;11(1). doi:10.1038/s41598-020-80756-6
38. Norcross JP, Lester GE, Weinhold P, Dahners LE. An in vivo model of degenerative disc disease. J Orthop Res. 2003;21(1):183–188. doi:10.1016/S0736-0266(02)00098-0
39. Urban JP, Roberts S. Degeneration of the intervertebral disc. Arthritis Res Ther. 2003;5:1–11. doi:10.1186/ar629
40. Zhou C, Yao S, Fu F, et al. Morroniside attenuates nucleus pulposus cell senescence to alleviate intervertebral disc degeneration via inhibiting ROS-Hippo-p53 pathway. Front Pharmacol. 2022;13:942435. doi:10.3389/fphar.2022.942435
41. Zhang A, Zheng J, Qin X, Yu N, Wang K-N. Recent Progress in Pyroptosis Probes and Inducers. Advanc Sens Energ Mat. 2024;3:100092. doi:10.1016/j.asems.2024.100092
42. Gordon C, Isenberg D. Systemic lupus Erythematosus. Oxford University Press; 2016.
43. Schile A, Petrillo M, Vovk A, et al. A comprehensive phenotyping program for the MRL-lpr mouse lupus model. J Immunol. 2018;200(1_Supplement):40. doi:10.4049/jimmunol.200.Supp.40.2
44. Andrews BS, Eisenberg RA, Theofilopoulos A, et al. Spontaneous murine lupus-like syndromes. Clinical and immunopathological manifestations in several strains. J Exp Med. 1978;148(5):1198–1215. doi:10.1084/jem.148.5.1198
45. Long Z, Zeng L, He Q, et al. Research progress on the clinical application and mechanism of iguratimod in the treatment of autoimmune diseases and rheumatic diseases. Front Immunol. 2023;14:1150661. doi:10.3389/fimmu.2023.1150661
46. Dent EL, Taylor EB, Turbeville HR, Ryan MJ. Curcumin attenuates autoimmunity and renal injury in an experimental model of systemic lupus erythematosus. Physiological Reports. 2020;8(13):e14501. doi:10.14814/phy2.14501
47. Munoz LE, van Bavel C, Franz S, Berden J, Herrmann M, Van Der Vlag J. Apoptosis in the pathogenesis of systemic lupus erythematosus. Lupus. 2008;17(5):371–375. doi:10.1177/0961203308089990
48. Sheriff A, Gaipl US, Voll RE, Kalden JR, Herrmann M. Apoptosis and systemic lupus erythematosus. Rheumatic Dis Clin. 2004;30(3):505–527. doi:10.1016/j.rdc.2004.04.006
49. Mende R, Vincent FB, Kandane-Rathnayake R, et al. Analysis of serum interleukin (IL)-1β and IL-18 in systemic lupus erythematosus. Front Immunol. 2018;9:1250. doi:10.3389/fimmu.2018.01250
50. Chen Y, Zhou C, Bian Y, et al. Cadmium exposure promotes thyroid pyroptosis and endocrine dysfunction by inhibiting Nrf2/Keap1 signaling. Ecotoxicol Environ Saf. 2023;249:114376. doi:10.1016/j.ecoenv.2022.114376
51. Yu P, Zhang X, Liu N, Tang L, Peng C, Chen X. Pyroptosis: mechanisms and diseases. Signal Transduc Target Therap. 2021;6(1):128. doi:10.1038/s41392-021-00507-5
52. Li Z, Guo J, Bi L. Role of the NLRP3 inflammasome in autoimmune diseases. Biomed Pharmacother. 2020;130:110542. doi:10.1016/j.biopha.2020.110542
53. Chen S, Jiang F, Ren J, Liu J, Meng W. Association of IL-18 polymorphisms with rheumatoid arthritis and systemic lupus erythematosus in Asian populations: a meta-analysis. BMC Med Genet. 2012;13:1–7. doi:10.1186/1471-2350-13-107
54. da Cruz HLA, Cavalcanti CAJ, de Azêvedo Silva J, et al. Differential expression of the inflammasome complex genes in systemic lupus erythematosus. Immunogenetics. 2020;72:217–224. doi:10.1007/s00251-020-01158-6
55. Zou H, Chen M, Wang X, et al. C/EBPβ isoform‐specific regulation of podocyte pyroptosis in lupus nephritis‐induced renal injury. J Pathol. 2023;261(3):269–285. doi:10.1002/path.6174
56. Kahlenberg JM, Carmona-Rivera C, Smith CK, Kaplan MJ. Neutrophil extracellular trap–associated protein activation of the NLRP3 inflammasome is enhanced in lupus macrophages. J Immunol. 2013;190(3):1217–1226. doi:10.4049/jimmunol.1202388
57. Xin Y, Gao C, Wang L, Liu Q, Lu Q. Lipopolysaccharide released from gut activates pyroptosis of macrophages via Caspase 11‐Gasdermin D pathway in systemic lupus erythematosus. MedComm. 2024;5(6):e610. doi:10.1002/mco2.610
58. Xie Y, Li X, Deng W, et al. Knockdown of USF2 inhibits pyroptosis of podocytes and attenuates kidney injury in lupus nephritis. J Molecul Histol. 2023;54(4):313–327. doi:10.1007/s10735-023-10135-8
59. Zhang C, Zhang C, Huang C, Ji J, Liu J, Lu Y. Hsa_circ_0012919 promotes pyroptosis in CD4+ T cells of systemic lupus erythematous by miR‐125a‐3p/GSDMD axis. Exp Dermatol. 2023;32(1):41–49. doi:10.1111/exd.14680
60. Liu Y, Lei H, Zhang W, et al. Pyroptosis in renal inflammation and fibrosis: current knowledge and clinical significance. Cell Death Dis. 2023;14(7):472. doi:10.1038/s41419-023-06005-6
61. S-M K, Lin J-C, Lin T-J, et al. Citral alleviates an accelerated and severe lupus nephritis model by inhibiting the activation signal of NLRP3 inflammasome and enhancing Nrf2 activation. Arthritis Res Ther. 2015;17:1–13. doi:10.1186/s13075-014-0514-0
62. Li M, Shi X, Qian T, et al. A20 overexpression alleviates pristine-induced lupus nephritis by inhibiting the NF-κB and NLRP3 inflammasome activation in macrophages of mice. Int J Clin Exp Med. 2015;8(10):17430.
63. Wang X, Zhou T, Yang X, et al. DDRGK1 enhances osteosarcoma chemoresistance via inhibiting KEAP1‐mediated NRF2 ubiquitination. Adv. Sci. 2023;10(14):2204438. doi:10.1002/advs.202204438
64. Barati MT, Caster DJ. The potential of Nrf2 activation as a therapeutic target in systemic lupus erythematosus. Metabolites. 2022;12(2):151. doi:10.3390/metabo12020151
65. Castejón ML, Alarcón-De-La-Lastra C, Rosillo MÁ, et al. A new peracetylated oleuropein derivative ameliorates joint inflammation and destruction in a murine collagen-induced arthritis model via activation of the Nrf-2/Ho-1 antioxidant pathway and suppression of MAPKs and NF-κB activation. Nutrients. 2021;13(2):311. doi:10.3390/nu13020311
66. Xiang Q, Zhao Y, Lin J, Jiang S, Li W. The Nrf2 antioxidant defense system in intervertebral disc degeneration: molecular insights. Exp Mol Med. 2022;54(8):1067–1075. doi:10.1038/s12276-022-00829-6
67. Shao Z, Wang B, Shi Y, et al. Senolytic agent Quercetin ameliorates intervertebral disc degeneration via the Nrf2/NF-κB axis. Osteoarthritis Cartilage. 2021;29(3):413–422. doi:10.1016/j.joca.2020.11.006
68. Liu Z, Bethunaickan R, Huang W, Ramanujam M, Madaio MP, Davidson A. IFN-α confers resistance of systemic lupus erythematosus nephritis to therapy in NZB/W F1 mice. J Immunol. 2011;187(3):1506–1513. doi:10.4049/jimmunol.1004142
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.