")
Back to Journals » Nature and Science of Sleep » Volume 17
Transcriptomic Changes and Pathological Mechanisms in Familial and Sporadic Idiopathic Restless Legs Syndrome: Implications for Inflammation and Cell Adhesion Molecules
Authors He XR , Song JM, Zhao JP, Zhang J, Feng JT, Chen SQ, Zhou ZY, Wang HM, Zhang Y, Feng Y, Wu YC, Zhu XY
Received 20 December 2024
Accepted for publication 9 May 2025
Published 11 June 2025 Volume 2025:17 Pages 1231—1247
DOI https://doi.org/10.2147/NSS.S512951
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sarah L Appleton
Xin-Rong He,1,* Jia-Min Song,1,* Jia-Peng Zhao,2 Jing Zhang,1 Jing-Tao Feng,1 Shu-Qin Chen,3 Zhi-Yuan Zhou,1 Hong-Ming Wang,4 Yue Zhang,5 Ya Feng,1 Yun-Cheng Wu,1 Xiao-Ying Zhu1
1Department of Neurology, Shanghai General Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, 200080, People’s Republic of China; 2Department of Orthopedics, Yongxiu County Traditional Chinese Medicine Hospital in Jiujiang City, Jiangxi, 330304, People’s Republic of China; 3Community Health Service Center, Ouyang Road District, Hongkou District, Shanghai, 200080, People’s Republic of China; 4Department of Clinical Laboratory Medicine, Shanghai General Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, 200080, People’s Republic of China; 5Department of Bioinformatics and Biostatistics, School of Life Sciences and Biotechnology, Shanghai Jiao Tong University, Shanghai, 200240, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yun-Cheng Wu, Department of Neurology, Shanghai General Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, 200080, People’s Republic of China, Tel +86-21-63240090, Email [email protected] Xiao-Ying Zhu, Department of Neurology, Shanghai General Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, 200080, People’s Republic of China, Email [email protected]
Purpose: Individuals affected by restless legs syndrome (RLS) tend to have familial predispositions without fully explained by genetic variants, and transcriptomic analysis may help elucidate the pathogenic mechanisms of RLS. The study aims to investigate transcriptomic changes and underlying pathological mechanisms in familial and sporadic idiopathic RLS to uncover potential contributors to its pathogenesis.
Patients and Methods: This study included 37 RLS patients, 39 unrelated healthy controls and 19 healthy relatives of RLS patients with a positive family history. Messenger RNA (mRNA) extracted from the peripheral blood mononuclear cells of these participants was analyzed via next-generation sequencing, followed by GO and KEGG pathway analyses. Differentially expressed mRNAs were validated by RT-qPCR in a subset of patients and controls. The relationships between the expression levels and clinical indices were evaluated via correlation analysis.
Results: After comparing with unrelated healthy controls and excluding genes with similar expression patterns in familial healthy controls, we identified nine upregulated and 28 downregulated mRNAs specifically in RLS patients. GO enrichment analysis indicated that these mRNAs are involved in protein binding and catalytic activity. KEGG analysis revealed that inflammation-related signaling pathways and cell adhesion molecules (CAMs) may be associated with RLS. Three specific mRNAs, including SPARCL1, CCL8 and SELE, demonstrated notably downregulated expression in RLS patients and were subsequently validated in a subset of 10 patients and 19 healthy controls.
Conclusion: This study revealed differentially expressed SPARCL1, CCL8 and SELE in RLS patients, indicating the potential involvement of inflammatory pathways and CAMs in RLS pathogenesis. These findings further support the association between RLS, inflammation, and synaptic transmission, providing insights into potential diagnostic and therapeutic strategies targeting these pathways.
Keywords: restless legs syndrome, peripheral blood mononuclear cells, bioinformatics, transcriptomics, inflammation, message-RNA sequencing
Introduction
Restless leg syndrome (RLS) is a neurological condition characterized by an uncontrollable urge to move the legs, typically accompanied by discomfort and unpleasant sensations in the lower extremities.1 The severity of symptoms typically follows a circadian pattern, worsening in the evening or at night and improving with movement of the lower limbs.1–4 Although peripheral iron metabolism disorders are common, only 10–20% of RLS patients present with decreased serum ferritin levels.5 Therapeutic options for RLS include iron supplements, dopamine agonists, calcium channel ligands and opioids.6,7 Notably, dopamine agonists can worsen the condition over time, a phenomenon known as augmentation, characterized by a gradual exacerbation of symptoms beyond pretreatment levels and the loss of the circadian pattern typically observed in RLS.8–10 Dopaminergic dysfunction and brain iron deficiency (BID) are widely believed to play key roles in the pathophysiology of RLS,11 along with the involvement of adenosinergic, GABAergic, opioid and glutamatergic neurotransmission systems.12 Additionally, inflammatory mechanisms have been implicated in RLS,13 as evidenced by elevated levels of inflammatory cytokines14 and changes in oxidative stress.15 However, the precise mechanisms remain to be further validated. Elucidating these pathways remains crucial for developing more effective therapies, particularly given RLS’s significant impact on sleep quality and quality of life.
Alterations in dopaminergic signaling are considered one of the primary pathological mechanisms of RLS, evidenced by symptom improvement with dopamine D2/3 receptor (D2/3R) agonists,16 and worsening with dopaminergic antagonists.17,18 However, the simplistic view of reduced DAergic function in RLS does not fully explain the circadian rhythm of RLS.19 Genetic studies have revealed significant associations between RLS and cell adhesion molecules (CAMs). Linkage studies in RLS families have identified at least eight susceptibility loci,20 and genome-wide association studies (GWASs) have so far identified 21 susceptibility genes.20–22 Among these genes, the myeloid ecotropic viral integration site one homolog (MEIS1), BTB domain containing 9 (BTBD9), and protein tyrosine phosphatase receptor type delta (PTPRD) genes are linked to dopaminergic transmission and iron metabolism.23 Notably, PTPRD24 may regulate synaptic states by mediating microcellular connectivity in dopaminergic neurons. Thus, revealing the circuits and synaptic connections involving CAMs is crucial. Over 60% of RLS patients report a positive family history, underscoring the significant role that genetic factors play in the etiology of this condition.25 Having a first-degree relative increases the risk of developing RLS by 6–7 times.25 Additionally, pathway analysis from a recent GWAS meta-analysis suggested a potentially pathological role for neurodevelopmental processes,22 which are essential for properly forming and maintaining functional neural circuits in the nervous system. Neurophysiological studies implicate that the dysfunction of complex cortico-subcortical networks may underlie RLS, highlighting the presence of hyperactive excitatory mechanisms and impaired inhibitory control in RLS.26,27 Such excitability alterations have been associated with altered sensory processing in RLS. Elena et al28 reported increased excitability in the primary somatosensory cortex (S1) and impaired somatosensory gating, which play a critical role in aberrant sensory information processing. Supporting this, deficits in cortical plasticity and sensorimotor integration have also been observed in RLS patients.29
Chronic inflammatory states and oxidative stress may contribute to the pathophysiology of RLS.30 Some studies have evaluated peripheral inflammatory markers in RLS patients and healthy controls31–34 and identified inflammatory cytokines and immune markers as potential biomarkers.15 Although proteomic studies have reported upregulation or downregulation of proteins involved in inflammatory processes, these findings should be considered preliminary.15 The potential role of inflammatory factors in RLS pathogenesis or their value as biomarkers remains to be determined.
Next-generation sequencing offers a powerful high-throughput approach to investigate transcriptomic alterations and associated signaling pathways in RLS patients. Several previous transcriptomic studies have provided valuable insights into RLS genetics and pathophysiology.33,35 Therefore, we hypothesize that differential mRNA expression patterns and associated signaling pathways in RLS patients, independent of familial genetic aggregation, may play a role in the pathogenesis of this disease. This could provide clues for new diagnostic and therapeutic approaches.
Materials and Methods
Subjects
From September 2022 to September 2023, we consecutively enrolled a total of 95 participants, including 37 RLS patients, 39 unrelated healthy controls (HCs) and 19 healthy relatives of RLS patients with a positive family history (familial healthy controls, fHC) from Shanghai General Hospital affiliated to Shanghai Jiao Tong University School of Medicine.
37 RLS patients were diagnosed by movement disorder specialists using International RLS Study Group (IRLSSG) criteria,36 exhibiting characteristic symptoms (urge to move/worsening at rest with lower-limb sensory disturbances relieved by movement), with an average disease duration of 13.0 years. As in our previous study,4 we conducted face‒to‒face interviews to collect the demographic information, disease duration, symptoms, medications, and findings from general neurological and medical examinations. Exclusion criteria included: (1) sleep disorders other than RLS; (2) psychiatric disorders; (3) neurological complications; or (4) use of any medications that could alter the circadian rhythm within three weeks before the study. All patients had no RLS comorbidities and were not taking dopaminergic or α2δ-ligand drugs.
Healthy controls were age-matched, medication-free, and had no history of sleep, physical, neurological, or psychiatric disorders, and primarily were the spouses of patients. 19 fHCs were used as a separate comparison group to explore potential familial or genetic contributions and excluded from the differential expression analysis between RLS patients and HCs. Detailed information on these RLS families is shown in Figure S1.
Peripheral iron status was evaluated by obtaining clinical laboratory data on TfR, ferritin, transferrin saturation (TSAT), and serum iron levels. Written informed consent was obtained from all participants, including those in the pedigree tree. This study was conducted under the principles of the Declaration of Helsinki and was approved by the Ethics Committee of Shanghai General Hospital affiliated to Shanghai Jiao Tong University School of Medicine (Ethics number: 2017–176).
Isolation of Cells, Total RNA Extraction and Quality Control
Total RNA extraction process included blood sample collection after assessments, PBMC isolation within 30 minutes, total RNA extraction and RNA storage. Venous blood was collected in the morning after all clinical assessments after a 12-hour fasting period. Blood samples were drawn into EDTA anticoagulant tubes and promptly transported on ice to the laboratory within 30 minutes to preserve cell viability. PBMCs were isolated via density gradient centrifugation via Ficoll‒Paque (GE Healthcare, USA), and the intermediate layer was extracted. Red blood cell lysis (Sigma‒Aldrich, USA) buffer was used at room temperature for five minutes, and the mixture was centrifuged at 250 ×g and 4°C for 5 min to remove any residual erythrocytes in the cell pellet. TRIzol reagent and DNase (Cat R401-01, Vazyme, Nanjing, China) were used to extract total RNA and ensure purity and accurate quantification according to the manufacturer’s protocol. The same procedures were followed for sample collection, storage, extraction and purification. RNA extraction for PBMC samples in the same group was performed simultaneously and completed within 7 days to ensure meaningful transcriptomic changes. The RNA was stored at −80°C, and its quality was evaluated via a NanoDrop™ 2000 spectrophotometer (Thermo Fisher Scientific, USA). Only samples with a 260/280 absorbance ratio between 1.8 and 2 were considered acceptable for further analysis.
Sequencing Library Construction
MRNA was extracted from total RNA for each group within 7 days using VAHTS® mRNA Capture Beads (N401-02, Human, Vazyme). Double-stranded cDNA was synthesized from purified polyadenylated mRNA templates using the PrimeScript™ Double Strand cDNA Synthesis Kit (TaKaRa), following the manufacturer’s protocol. cDNA libraries for Illumina sequencing were prepared using the VAHTS Universal V6 RNA-seq Library Prep Kit for Illumina® (NR604-02, Vazyme). The library concentration was quantified using a Qubit 2.0 Fluorometer (Thermo Fisher Scientific, USA), and library fragment distribution was assessed with an Agilent 4200 TapeStation (Agilent Technologies, China).
Transcriptomic and Functional Enrichment Analysis
Before sequencing, the concentration and size distribution of the cDNA library were assessed using a Qubit 2.0 fluorometer (Thermo Fisher Scientific) and an Agilent 4200 TapeStation (Agilent Technologies) to ensure RNA integrity (RIN ≥ 9). Sequencing was then performed on an Illumina NovaSeq 6000 platform, generating raw image files that were converted into reads. To improve data quality, low-quality reads were filtered out using Seqtk (v1.0) based on the following criteria: (1) removal of adapter sequences, (2) trimming of bases with a quality score below 20 at the 3′ end, (3) exclusion of reads shorter than 25 nucleotides, and (4) removal of ribosomal RNA reads, resulting in a set of clean reads. The spliced mapping algorithm of Hisat2 (v2.0.4) was applied to perform Genome mapping on the preprocessed reads. To ensure comparability of gene expression levels across different genes and samples, read counts were normalized to fragments per kilobase of transcript per million mapped reads (FPKM) values, following the method described in Mapping and quantifying mammalian transcriptomes by RNA-Seq. Gene fragments were counted using StringTie (v1.3.0) after alignment with HISAT2, and normalization was performed using the trimmed mean of M values (TMM) method, as described in A scaling normalization method for differential expression analysis of RNA-seq data. Sample relationships were assessed through correlation analysis and principal component analysis (PCA), with batch effects corrected accordingly. Differential gene expression analysis was conducted using edgeR (v3.2.0), with age and sex included as covariates. Q value was corrected after false discovery rate (FDR) correction applied to control the P-value threshold, and fold-change values were calculated to identify differentially expressed genes. Differentially expressed mRNAs were defined as those identified with a q value ≤ 0.05 and a fold change (FC) ≥ 1.5. The average level of mRNAs was calculated via the average expression function. The selected differentially expressed genes were mapped to GO and KEGG terms. Terms with a q-value ≤ 0.05 after multiple hypothesis correction (FDR correction) were considered significantly enriched. GO enrichment analysis, KEGG enrichment analysis, and visualization were performed via the ClusterProfiler (v4.0.0) R package.
Preparation of cDNA and Real-Time Quantitative PCR Analysis
cDNA synthesis was conducted with 200 ng of total RNA via a TB Green Premix Ex Taq II (Tli RNaseH Plus) Kit (Cat RR820B, TaKaRa, Dalian, China) following the manufacturer’s protocol. Real-time quantitative PCR (RT-qPCR) was conducted via the LightCycler® 480 System (Roche, Basel, Switzerland) to assess the mRNA expression levels. Three technical and biological replicates were performed to verify PCR efficiency (90–110%), and melting curve analysis was checked for non-specific amplification. The primer sequences were as follows: for SPARC-like protein 1 (SPARCL1), forward: 5′- ACGGTAGCACCTGACAACAC-3′ and reverse: 5′- ATGGTGGGAATCGTCTTCTGT-3; for proinflammatory C–C motif chemokine ligand 2 (CCL2), forward: 5′- CAGCCAGATGCAATCAATGCC-3′ and reverse: 5′- TGGAATCCTGAACCCACTTCT-3′; for proinflammatory C–C motif chemokine ligand 8 (CCL8), forward: 5′- TGGAGAGCTACACAAGAATCACC-3′ and reverse: 5′- TGGTCCAGATGCTTCATGGAA-3′; for endothelial adhesion molecule 1 (SELE), forward: 5′- CAGCAAAGGTACACACACCTG-3′ and reverse: 5′- CAGACCCACACATTGTTGACTT-3′; for Homo sapiens apolipoprotein A-II (APOA2), forward: 5′- CTGTGCTACTCCTCACCATCT-3′ and reverse: 5′- CTCTCCACACATGGCTCCTTT-3′. Target gene expression levels were calculated compared to housekeeping gene 18S rRNA (forward: 5′-GCAATTATTCCCCATGAACG-3′ and reverse: 5′-GGGACTTAATCAACGCAAGC-3′). Relative mRNA expression levels were described using the 2 − ΔΔCt method (ΔCt = Cttarget − Ctreference, − ΔΔCt = sample ΔCT − 18S ΔCT).
Statistical Analysis
Quantitative variables are described as either the median with interquartile range (25th–75th percentiles) or the mean ± standard deviation (SD). Group comparisons were performed using Welch’s t-test or Mann–Whitney U-test. For RNA sequencing data, differential gene expression analysis was performed using edgeR, and p-values were adjusted by FDR correction (q-values). Genes with q < 0.05 and fold-change (FPKM-based) were considered significant. Spearman correlation corrected for age was used to test the association between levels of mRNA levels and clinical index. Statistical significance was defined as *P < 0.05; **P< 0.01; ***P< 0.001; ****P < 0.001; ns, not significant. SPSS version 20.0 (IBM Corporation, USA) was utilized to conduct statistical analyses, and Prism software (version 8.0; GraphPad Software, USA) was used for data visualization.
Result
Clinical Characteristics of the Participants
During the screening phase, we recruited 37 RLS patients, 39 matched HCs, and 19 familial healthy controls (fHCs) to minimize the impact of familial genetic susceptibility (Figure S1). In the validation phase, a subset of 10 RLS patients and 19 HCs were included and matched by sex and age. The demographic and disease characteristics of all participants are summarized in Table 1. Notably, serum iron levels were significantly lower in RLS patients than in HCs (P = 0.0198).
 |
Table 1 Demographic and Clinical Characteristics of Participants |
mRNA Expression Profiles of RLS Patients and Healthy Controls
The research design is shown in Figure 1A. Hierarchical cluster analysis performed using the PBMC mRNA results revealed significant differences in mRNA expression between RLS patients and HCs (Figure 1B). In total, 71 differentially expressed mRNAs were identified, with 18 significantly elevated and 53 significantly reduced in RLS patients (q value < 0.05) (Figure 1C; the detailed information is listed in Table S1). Among them, MTND1P23 exhibited the highest upregulation with a fold change of 5.6, while genes such as APOA2 and SPARCL1 showed the most significant downregulation (each with a fold change below −5) (Table S1).
 |
Figure 1 Transcriptomic analysis of RLS patients and healthy controls. (A) Study design and timeline. (B) Heatmap illustrating the differential expression levels of mRNAs identified in restless legs syndrome (RLS) patients and healthy controls (HCs). The X-axis represents the patient and control groups from the cluster analysis, whereas the Y-axis lists the 71 differentially expressed mRNAs. Red and green indicate upregulation and downregulation, respectively. (C) Volcano plot depicting the differences in mRNA expression between patients and HCs. The X-axis represents the log2 (fold change) values, whereas the Y-axis represents the -log10 (q value) values. The gray dots indicate mRNAs without significant expression differences, the red dots represent mRNAs with upregulated expression, and the blue dots indicate mRNAs with downregulated expression. Differentially expressed genes were identified using edgeR, with a false discovery rate (FDR)-adjusted q value ≤ 0.05 and an absolute log2 fold change (|log2FC|) ≥ 1.5. Abbreviations: RLS, restless legs syndrome; HC, healthy control; FDR, false discovery rate. |
Functional and Pathway Enrichment Analysis of Differentially Expressed mRNAs
To elucidate the biological roles of the differentially expressed mRNAs, GO and KEGG analyses were performed on 71 genes (q value ≤ 0.05 and |FC| ≥ 1.25) (Figure 2A and B). GO enrichment analysis indicated that these mRNAs were associated with various molecular functions, biological processes, and cellular components (Tables S2–S4). Notably, upregulated genes such as MTCO1P40, carbonic anhydrase 1 (CA1), CBY2, and ubiquitin-associated protein 2 (UBBP2) were found to be associated with the following molecular functions: binding, protein binding and catalytic activity. Among the downregulated genes, APOA2 and SPARCL1 were involved in binding, protein binding, and ion binding. KEGG analysis identified 22 signaling pathways, with the q value threshold set at 0.05, among which the interleukin 17 (IL-17) signaling pathway, tumor necrosis factor (TNF) signaling pathway and CAMs were the most prominent. The differentially expressed mRNAs involved in each pathway, along with the distribution of these pathways across various processes, functions, or organs, are presented in Table S5. Additionally, functional enrichment analysis was performed separately for upregulated and downregulated genes. GO analysis revealed that upregulated genes were primarily enriched in immune response pathways (Figure 2C), while downregulated genes were mainly associated with the regulation of leukocyte migration and humoral immune response (Figure 2D). KEGG analysis further showed that downregulated genes were enriched in cytoskeleton-related pathways in muscle cells (Figure 2E and F).
 |
Figure 2 GO and KEGG analysis of differentially expressed mRNAs in RLS patients. (A) Bubble plot illustrating selected enriched GO terms. The color gradient represents q values, with red indicating smaller q values. Larger bubbles correspond to a greater number of genes involved in each pathway. Different shapes represent molecular functions (MF), biological processes (BP), and cellular components (CC). (B) Bubble plot displaying selected enriched KEGG pathways. The color gradient reflects the q values, with red signifying smaller q values. Larger bubbles indicate that more genes are involved in each pathway. (C) GO analyses of upregulated genes. (D) GO analyses of downregulated genes. (E)KEGG analyses of upregulated genes. (F) KEGG analyses of downregulated genes. For (C–F), the bubble size and color gradient follow the same conventions as in (A and B). GO and KEGG pathway enrichment analyses were performed based on differentially expressed genes (DEGs). P-values were corrected for multiple hypothesis testing using the false discovery rate (FDR) method, with a significance threshold set at q-value ≤ 0.05. GO terms and KEGG pathways meeting this criterion were considered significantly enriched. Abbreviations: RLS, restless legs syndrome; GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; MF, molecular function; BP, biological process; CC, cellular component; DEG, differentially expressed gene; FDR, false discovery rate. |
mRNA Expression Profiles of Related Familial Healthy Controls
To further investigate the distinct differences in mRNA expression in RLS patients, we compared the mRNA expression profiles of 19 fHC and 39 HCs. In the fHC group, we identified 247 mRNAs with upregulated expression (q < 0.05) and 894 with downregulated expression (q < 0.05) (additional data are listed in Table S6). To minimize the influence of familial genetic aggregation, we filtered out those mRNAs with similar up- or downregulation trends in both the RLS patient and fHC groups. Among them, nine mRNAs were consistently upregulated, and 25 were downregulated in both RLS patients and fHCs (Table S7). Notably, PTN exhibited an increasing trend across the RLS, fHCs, and HCs groups in the sequencing data (Figure S2). The specific and shared DEGs were demonstrated in Figure 3A and B. This process allowed us to focus on the mRNAs demonstrating uniquely upregulated (9 mRNAs) or downregulated (28 mRNAs) expression in RLS patients (q < 0.05, Table 2).
 |
Table 2 37 mRNAs Specifically Differentially Expressed in RLS Patients Compared to Both fHCs and HCs |
 |
Figure 3 Venn diagram of specific and shared DEGs between RLS patient and fHC. (A) Upregulated DEGs in RLS patients and fHCs. The numbers indicate unique and shared DEGs between the two groups. (B) Downregulated DEGs in RLS patients and fHCs. The numbers in each section represent the DEGs that are specific to RLS patients, specific to fHCs, or shared between the two groups. Statistical significance determined by an adjusted q-value ≤ 0.05 and |log2FC| ≥ 1. Abbreviations: DEGs, differentially expressed genes; RLS, restless legs syndrome; fHC, familial healthy controls. |
Validation of Candidate mRNAs Screened by RT-qPCR
We selected SPARCL1, CCL2, CCL8, SELE and APOA2 from the 37 significantly downregulated mRNAs in RLS patients. The expression levels of these genes were validated in a subset of 10 RLS patients and 19 HCs via RT-qPCR (Table 3). The results confirmed significantly lower levels of SPARCL1, CCL8 and SELE in RLS patients compared to HCs (Figure 4A, C and D), while CCL2 and APOA2 showed no significant differences (Figure 4B and E).
 |
Table 3 Differences in the Expression of the 4 Candidate mRNAs in the Validation Cohort (N = 29) |
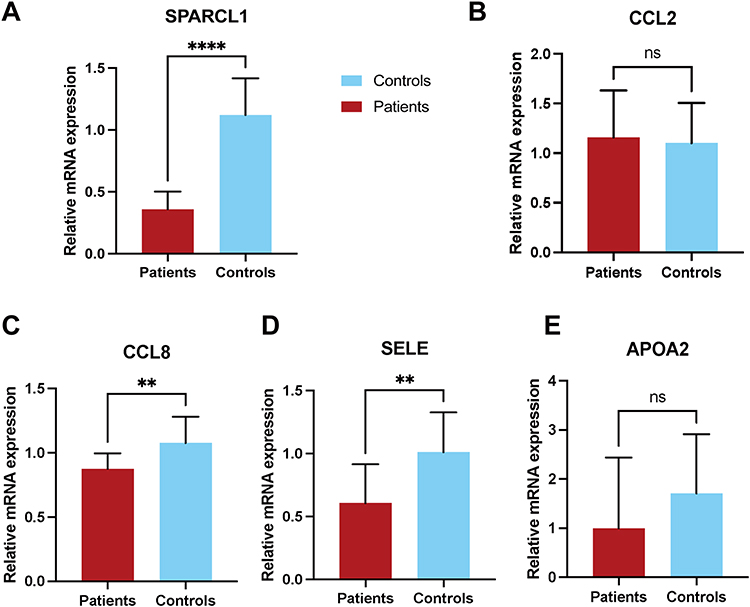 |
Figure 4 Validation of differential expression of five candidate mRNAs in an independent cohort. (A)Relative expression levels of SPARCL1 were compared between RLS patients and HCs. (B) Relative expression levels of CCL2 were compared between RLS patients and HCs. (C) Relative expression levels of CCL8 were compared between RLS patients and HCs. (D) Relative expression levels of SELE were compared between RLS patients and HCs. (E) Relative expression levels of APOA2 were compared between RLS patients and HCs. Statistical significance was determined using an unpaired two-tailed t-test. Data are presented as mean ± standard deviation. **P < 0.01; ****P < 0.0001; ns, not significant. Abbreviations: RLS, restless legs syndrome; HC, healthy controls; SPARCL1, SPARC-like protein 1; CCL2, proinflammatory C–C motif chemokine ligand 2; CCL8, proinflammatory C–C motif chemokine ligand 8; SELE, endothelial adhesion molecule 1; APOA2, Homo sapiens apolipoprotein A-II. |
Relationships between Five Selected mRNAs and Clinical Indices
Correlation analysis of five selected mRNAs, including SPARCL1, CCL2, CCL8, APOA2 and SELE in patients, as presented in Table 4, revealed no significant associations between their downregulated expression and RLS severity, as assessed by IRLSS or peripheral iron deficiency-related laboratory parameters (TfR, ferritin, TSAT, and iron).
 |
Table 4 Correlation Analysis of Four Candidate mRNAs with Clinical Indicators |
Discussion
In this study, we investigated the transcriptomic characteristics of the mRNA expression profiles in RLS patients’ PBMCs. A total of 71 candidate mRNAs were identified by comparing RLS patients and HCs. After excluding the influence of familial aggregation, we identified 9 upregulated and 28 downregulated mRNAs specific to RLS patients. RT-qPCR confirmed significant downregulation of SPARCL1, CCL8, and SELE in RLS PBMCs. GO and KEGG analyses indicated potential involvement of the IL-17 and TNF signaling pathways in the pathogenesis of RLS. While these findings suggest that inflammation and immunity may be important contributing factors to the pathophysiology of RLS, further research is needed to confirm these associations and elucidate the underlying mechanisms.
Among genes with a consistent upregulation trend between RLS patients and fHCs, PTN showed an increasing trend across RLS, fHCs, and HCs, after excluding pseudogenes and novel protein-encoding genes. PTN is known for its role in promoting neurite outgrowth.37 Although this trend was not successfully validated, its dysregulation may still represent a potential genetic risk factor for RLS progression.
Inflammation and Immunity
Our research findings suggest that inflammatory mechanisms are involved in RLS, as indicated by the involvement of the TNF and IL-17 signaling pathways. This aligns with a previous transcriptomic study supporting the involvement of inflammatory network mechanisms and immune processes in RLS. That study identified IL-17, chemokines, and inflammatory mediators of TRP channels, further suggesting that these mediators may be associated with sensory symptoms in RLS patients.33 Proteomic studies in RLS patients have revealed elevated levels of reactive oxygen species, activation of complement components,15 and increased levels of inflammatory proteins,14 further highlighting the role of inflammatory mechanisms. Previous studies have reported significantly higher levels of serum or plasma tumor necrosis factor α (TNF-α), interleukin-1β (IL-1β) and interleukin-6 (IL-6) in RLS patients than in controls, indicating systemic inflammatory changes.38 Our findings reinforce the involvement of the TNF signaling pathway in the pathogenesis of RLS,31,38,39 suggesting a potential possible link between RLS symptoms and comorbid psychosomatic disorders.39 Interestingly, Trotti et al40 reported that among 137 untreated RLS patients, those with frequent periodic limb movements in sleep (PLMS) were more likely to have elevated levels of hypersensitive C-reactive protein (hs-CRP), although the IL-6 and TNF-α levels were not significantly increased.
Increasing evidence indicates that an abnormal immune regulatory response contributes to RLS, as shown by its higher prevalence in multiple sclerosis (MS)41 and rheumatoid arthritis (RA).42 Additionally, an RNA transcriptomic study has implicated the role of inflammatory cytokines in Parkinson’s disease (PD) pathogenesis, further supporting a potential immune-RLS link, given RLS is a common comorbidity in patients with PD.43 We observed an enrichment of differentially expressed mRNAs in the IL-17 pathway in RLS patients, further emphasizing the contribution of immune mechanisms to the pathogenesis of RLS. Autopsy studies on RLS patients showed microvascular T-cell infiltration,44,45 which may mediate the inflammatory infiltration of microvessels through mechanisms involving leukocyte transendothelial migration. Additionally, alterations in IL-17 may also be associated with RLS by affecting impaired dopamine synthesis, leading to altered dopaminergic signaling.46 We also detected the downregulation of CCL2 and CCL8 expression in RLS, which may play a role in the immunomodulatory function of sensory neurons. The anti-inflammatory molecule interleukin 10 (IL-10) has been reported to bind to receptors expressed on somatosensory neurons on the dorsal root ganglion (DRG)47 and trigeminal ganglion (TG),48 thereby downregulating CCL2.49 Given that DRG neuron-derived CCL2 is involved in immune cell infiltration and activation,50 the restriction of the ability of somatosensory neurons to respond to these alterations through immune modulation is reasonable. Our findings increase the understanding of neuroimmunity in regulating important sensory symptoms in RLS. In conclusion, our findings strengthen the evidence linking inflammation and immunity to RLS. However, the specific functions of inflammatory and immune factors in the pathogenesis and their potential as biomarkers require further investigation in future studies.
Neurotransmission and Synapse
Dopamine dysfunction in RLS is characterized by dysregulation, including increased presynaptic DA synthesis and release, decreased DA uptake, and reduced postsynaptic receptor density.19 This alteration in the dynamic relationship between presynaptic and postsynaptic functions involves complex changes in multiple elements. One perspective is that differences in microscopic intercellular connectivity among dopaminergic neurons may explain these changes.51 Pathway analysis of the differentially expressed mRNAs suggests that CAMs play a role. CAMs are described as potential candidates for playing a central role in determining synaptic connectivity and neural development.52 PTPRD, a CAM, has been identified repeatedly associated with RLS in several GWAS.53–55 Among these studies, single-nucleotide polymorphisms (SNPs) in PTPRD are associated with reduced mRNA expression.56,57 Axonal PTPRD is involved in motor neuron axon termination and synapse formation during embryonic development58 via postsynaptic ligand interactions.59–65 PTPRD knockout mice exhibit impaired neuronal differentiation and cortical disruption, underscoring its role in neural connectivity.66 Notably, PTPRD is highly expressed in dopaminergic neurons, suggesting its involvement in dopaminergic neural circuit remodeling. Although whether PTPRD mutations directly impair dopaminergic neurotransmission remains unclear, these findings support its critical role in the synaptic connectivity, potentially leading to altered susceptibility to RLS.24,67 Dopaminergic network abnormalities in RLS patients and worsening symptoms with prolonged dopamine agonist use may be linked to CAM-mediated circuit dysfunction.68 A detailed elaboration of CAM-regulated synaptic connections could clarify treatment-related complications and identify new therapeutic targets.
A novel finding in our study was the downregulation of SPARCL1 in RLS patients. SPARC, a matricellular protein (MCP), modulates tissue development and remodeling modulator by influencing key cellular processes such as cell adhesion and proliferation.69 SPARCL1, expressed primarily in the central nervous system, is reported to be associated with the formation and maturation of excitatory synapses.70,71 Specifically, SPARCL1 facilitates the development of synaptic N-methyl-d-aspartate (NMDA) receptors, which can be essential for synaptic connectivity.72 Clinical evidence suggests the involvement of NMDA receptor-mediated alterations in glutamatergic neurotransmission in RLS pathogenesis. Noncompetitive NMDA receptor inhibitors, such as ketamine, have shown efficacy in alleviating RLS symptoms,73 and another NMDA antagonist, antamethadone, has also proven effective.74 Although a direct link between SPARCL1 and RLS has not yet been established, our results may suggest the potential role of SPARCL1 in NMDA receptor-mediated glutamatergic dysregulation in RLS. A recent optogenetic study in rats with BID demonstrated hypersensitivity in corticostriatal glutamatergic terminals.75 This hyperglutamatergic state may be related to the hyperarousal often observed in RLS patients.76 Given that BID is prevalent in the majority of RLS patients, our results could provide valuable insights into SPARCL1’s role in synaptic adaptation and its potential contribution to the hyperarousal state in RLS.
Additionally, NMDA receptor-mediated glutamatergic neurotransmission may also be associated with maladaptive synaptic plasticity in the brain,77,78 which could explain RLS-related sensory symptoms such as chronic pain. A study of neuropathic pain in SPARCL1 knockout mice suggested that SPARCL1-mediated enhancement of NMDA currents plays a key role in chronic pain maintenance.75 In particular, sensory neurons expressing SPARCL1 have been identified in human sensory ganglia, indicating the participation of SPARCL1 in the transmission of nociceptive signals.79 Therefore, one hypothesis is that SPARCL1 may modulate synaptic modifications to regulate the transsynaptic transmission of nociceptive signals, potentially alleviating the discomfort associated with sensory symptoms in RLS. The possible role of SPARCL1 in sensory adaptations is a perspective that has not been previously addressed. However, given the scarcity of relevant studies, the role of SPARCL1 in RLS needs to be further investigated. These observations are clinically significant, as inhibitors targeting inflammatory pathways and synaptic regulation may be promising therapeutic options for RLS.
Although we implemented strict inclusion and exclusion criteria, certain conditions have not been extensively explored concerning RLS, such as nonanemic iron deficiency80 and undiagnosed cardiovascular abnormalities.81 Residual effects of prior medications (eg, immunosuppressants/antidepressants) may have influenced the observed gene expression variability, although none of the participants were actively taking dopaminergic or α2δ-ligand medications. While our findings provide transcriptional insights, limitations include the lack of protein-level validation, a relatively small sample size, and potential discrepancies between peripheral blood transcriptomics and CNS pathology. Expanding the cohort size and incorporating more comprehensive clinical assessments, such as sleep parameters, disease severity, and peripheral iron status, are needed to clarify potential correlations and enhance the observations of our findings.
Conclusion
To summarize, the pathophysiological mechanisms of RLS are intricate and multifaceted. Our bioinformatics analysis identified 37 differentially expressed mRNAs (9 upregulated, 28 downregulated) in RLS patients after accounting for familial genetic aggregation. Notably, we validated significant downregulation of SPARCL1, CCL8 and SELE, whereas no significant difference was observed in CCL2 and APOA2 levels. Correlation analysis did not reveal significant associations between the expression of these five mRNAs and RLS severity or peripheral iron status. Functional analyses implicated the IL-17 and TNF signaling pathways as well as CAMs in RLS pathogenesis, suggesting two potential therapeutic avenues: anti-inflammatory therapies and synaptic intervention targeting connectivity in RLS management. These findings reinforce the growing evidence for inflammatory and synaptic adhesion mechanisms in RLS while highlighting the need for functional validation of identified pathways, RLS subtyping based on transcriptomic profiles and development of precision medicine strategies.
Abbreviations
RLS, restless legs syndrome; mRNA, Messenger RNA; PBMCs, peripheral blood mononuclear cells; NGS, next-generation sequencing; KEGG, Kyoto Encyclopedia of Genes and Genomes; RT-qPCR, real-time quantitative polymerase chain reaction; CAMs, cell adhesion molecules; BID, brain iron deficiency; PTPRD, protein tyrosine phosphatase receptor type delta; TfR, transferrin receptors; GWAS, genome-wide association studies; MEIS1, myeloid ecotropic viral integration site 1 homolog; BTBD9, BTB domain containing 9; HC, healthy controls; fHC, healthy relatives of RLS patients with a family history; TSAT, transferrin saturation; SPARCL1, SPARC-like protein 1; CCL2, pro-inflammatory C–C motif chemokine ligand 2; CCL8, pro-inflammatory C–C motif chemokine ligand 8; SELE, endothelial adhesion molecule 1; MTND1P23, Mitochondrially Encoded NADH Dehydrogenase 1 Pseudogene 23; MTCO1P40, Mitochondrially Encoded Cytochrome C Oxidase I Pseudogene 40; CBY2, chibby family member 2; POC1B-GALNT4, polypeptide N-acetylgalactosaminyltransferase 4; PLP1, proteolipid protein 1; STOX2, storkhead box 2; RPL7AP28, Ribosomal Protein L7/L12 Family Member AP28; APOA2, apolipoprotein A-II; BLOC1S5-TXNDC5, BLOC1S5-TXNDC5 readthrough, NMD candidate; TUBB8P7, Tubulin beta 8 class VIII; CA1, Carbonic anhydrase 1; UBBP2, Ubiquitin Associated Protein 2; IL-17, interleukin 17; TNF, tumor necrosis factor; TNF-α, tumor necrosis factor α; IL-1β, interleukin-1β; IL-6, interleukin-6; PLMS, periodic limb movements in sleep; Hs-CRP, hypersensitive C-reactive protein; MS, multiple sclerosis; RA, rheumatoid arthritis; IL-10, interleukin 10; DRG, dorsal root ganglion; TG, trigeminal ganglion; SNPs, single nucleotide polymorphisms; MCP, matricellular protein; NMDA, N-methyl-d-aspartate.
Data Sharing Statement
The data that support the findings of this study are available on request from the corresponding author, Xiaoying Zhu (Shanghai General Hospital, Shanghai Jiao Tong University School of Medicine, China). The data are not publicly available due to information that could compromise the privacy of research participants.
Ethics Statement and Informed Consent
The research was approved by the Ethics Committee of Shanghai General Hospital affiliated to Shanghai Jiao Tong University School of Medicine according to the “1964 Declaration of Helsinki”, and all participants provided written informed consent to participate in the study. In addition, written informed consent for publication was obtained from all individuals included in the pedigree tree and other relevant participants.
Acknowledgments
We appreciate all the patients for their participation in the study.
Author Contributions
All authors contributed substantially to the research, including the conceptualization, design, implementation, data collection, analysis, and interpretation of the study. They were involved in drafting, revising, and critically reviewing the manuscript. Each author provided final approval for the version to be published, agreed on the journal for submission, and accepted responsibility for the article’s content. The individual contributions of the authors, based on the CRediT taxonomy, are as follows:
Xin-Rong He: Conceptualization, Methodology, Data Curation, Writing - Original Draft Preparation, Writing - Review & Editing, Supervision, Software, Visualization. Jia-Min Song: Writing - Review & Editing, Methodology, Investigation, Data Collection, Software, Formal Analysis. Jia-Peng Zhao: Data Collection, Formal Analysis, Visualization, Writing - Review & Editing. Jing Zhang: Visualization, Investigation, Writing - Review & Editing. Jing-Tao Feng: Software, Validation, Writing - Review & Editing. Shu-Qin Chen: Resources, Writing - Review & Editing. Zhi-Yuan Zhou: Resources, Investigation, Writing - Review & Editing. Hong-Ming Wang: Data Curation, Investigation, Writing - Review & Editing. Yue Zhang: Supervision, Project Administration, Writing - Review & Editing. Ya Feng: Supervision, Resources, Writing - Review & Editing. Yun-Cheng Wu: Formal Analysis, Visualization, Writing - Review & Editing. Xiao-Ying Zhu: Conceptualization, Supervision, Funding Acquisition, Writing - Review & Editing.
All authors agreed on the journal to which the article will be submitted, reviewed and agreed on all versions of the article before submission, during revision, the final version accepted for publication, and any significant changes introduced at the proofing stage, agreed to take responsibility and be accountable for the contents of the article.
Funding
This work was supported by the National Natural Science Foundation of China (grant number 81901344), Shanghai Municipal Planning Commission of Science and Research Fund (201740017) and the Clinical Research Innovation Plan of Shanghai General Hospital (CTCCR-2018C02) and the Ministry of Science and Technology of the People’s Republic of China (2021ZD0201900, 2021ZD0201905).
Disclosure
We have no conflicts of interest to disclose.
References
1. Hening WA, Walters AS, Wagner M, et al. Circadian rhythm of motor restlessness and sensory symptoms in the idiopathic restless legs syndrome. Sleep. 1999;22(7):901–912. doi:10.1093/sleep/22.7.901
2. Silva C, Peralta AR, Bentes C. The urge to move and breathe - the impact of obstructive sleep apnea syndrome treatment in patients with previously diagnosed, clinically significant restless legs syndrome. Sleep Med. 2017;38:17–20. doi:10.1016/j.sleep.2017.06.023
3. Stevens MS. Restless legs syndrome/Willis-ekbom disease morbidity: burden, quality of life, cardiovascular aspects, and sleep. Sleep Med Clin. 2015;10(3):369–373, xv–xvi. doi:10.1016/j.jsmc.2015.05.017
4. Zhu XY, Wu TT, Wang HM, et al. Clinical features and subtypes of restless legs syndrome in Chinese population: a study of 359 patients. Sleep Med. 2019;59:15–23. doi:10.1016/j.sleep.2019.01.053
5. Allen RP, Earley CJ. The role of iron in restless legs syndrome. Mov Disord. 2007;22(Suppl 18):S440–448. doi:10.1002/mds.21607
6. Allen RP, Picchietti DL, Auerbach M, et al. Evidence-based and consensus clinical practice guidelines for the iron treatment of restless legs syndrome/Willis-Ekbom disease in adults and children: an IRLSSG task force report. Sleep Med. 2018;41:27–44. doi:10.1016/j.sleep.2017.11.1126
7. Silber MH, Buchfuhrer MJ, Earley CJ, Koo BB, Manconi M, Winkelman JW. The management of restless legs syndrome: an updated algorithm. Mayo Clin Proc. 2021;96(7):1921–1937. doi:10.1016/j.mayocp.2020.12.026
8. Allen RP, Ondo WG, Ball E, et al. Restless legs syndrome (RLS) augmentation associated with dopamine agonist and levodopa usage in a community sample. Sleep Med. 2011;12(5):431–439. doi:10.1016/j.sleep.2011.03.003
9. Lipford MC, Silber MH. Long-term use of pramipexole in the management of restless legs syndrome. Sleep Med. 2012;13(10):1280–1285. doi:10.1016/j.sleep.2012.08.004
10. Mitterling T, Heidbreder A, Stefani A, et al. Natural course of restless legs syndrome/Willis-Ekbom disease: long-term observation of a large clinical cohort. Sleep Med. 2015;16(10):1252–1258. doi:10.1016/j.sleep.2015.05.028
11. Mogavero MP, Mezzapesa DM, Savarese M, DelRosso LM, Lanza G, Ferri R. Morphological analysis of the brain subcortical gray structures in restless legs syndrome. Sleep Med. 2021;88:74–80. doi:10.1016/j.sleep.2021.10.025
12. Jiménez-Jiménez FJ, Alonso-Navarro H, García-Martín E, Agúndez JAG. Neurochemical features of idiopathic restless legs syndrome. Sleep Med Rev. 2019;45:70–87. doi:10.1016/j.smrv.2019.03.006
13. Buratti L, Petrelli C, Viticchi G, Provinciali L, Silvestrini M. Severe recurrent restless legs syndrome associated with systemic inflammation. Sleep Med. 2017;29:90–91. doi:10.1016/j.sleep.2016.07.024
14. Shin JW, Lee JH, Kim H, et al. Bioinformatic analysis of proteomic data for iron, inflammation, and hypoxic pathways in restless legs syndrome. Sleep Med. 2020;75:448–455. doi:10.1016/j.sleep.2020.09.002
15. Mondello S, Kobeissy FH, Mechref Y, et al. Searching for novel candidate biomarkers of RLS in blood by proteomic analysis. Nat Sci Sleep. 2021;13:873–883. doi:10.2147/NSS.S311801
16. Hening WA, Allen RP, Earley CJ, Picchietti DL, Silber MH. An update on the dopaminergic treatment of restless legs syndrome and periodic limb movement disorder. Sleep. 2004;27(3):560–583. doi:10.1093/sleep/27.3.560
17. Farde L. Selective D1- and D2-dopamine receptor blockade both induces akathisia in humans--a PET study with [11C]SCH 23390 and [11C]raclopride. Psychopharmacology. 1992;107(1):23–29. doi:10.1007/BF02244961
18. Winkelmann J, Schadrack J, Wetter TC, Zieglgänsberger W, Trenkwalder C. Opioid and dopamine antagonist drug challenges in untreated restless legs syndrome patients. Sleep Med. 2001;2(1):57–61. doi:10.1016/S1389-9457(00)00025-3
19. Earley CJ, Connor J, Garcia-Borreguero D, et al. Altered brain iron homeostasis and dopaminergic function in restless legs syndrome (Willis-Ekbom Disease). Sleep Med. 2014;15(11):1288–1301. doi:10.1016/j.sleep.2014.05.009
20. Jiménez-Jiménez FJ, Alonso-Navarro H, García-Martín E, Agúndez JAG. Genetics of restless legs syndrome: an update. Sleep Med Rev. 2018;39:108–121. doi:10.1016/j.smrv.2017.08.002
21. Didriksen M, Nawaz MS, Dowsett J, et al. Large genome-wide association study identifies three novel risk variants for restless legs syndrome. Commun Biol. 2020;3(1):703. doi:10.1038/s42003-020-01430-1
22. Schormair B, Zhao C, Bell S, et al. Identification of novel risk loci for restless legs syndrome in genome-wide association studies in individuals of European ancestry: a meta-analysis. Lancet Neurol. 2017;16(11):898–907. doi:10.1016/S1474-4422(17)30327-7
23. Trenkwalder C, Allen R, Högl B, et al. Comorbidities, treatment, and pathophysiology in restless legs syndrome. Lancet Neurol. 2018;17(11):994–1005. doi:10.1016/S1474-4422(18)30311-9
24. Schormair B, Kemlink D, Roeske D, et al. PTPRD (protein tyrosine phosphatase receptor type delta) is associated with restless legs syndrome. Nat Genet. 2008;40(8):946–948. doi:10.1038/ng.190
25. Allen RP, La Buda MC, Becker P, Earley CJ. Family history study of the restless legs syndrome. Sleep Med. 2002;3(3 Suppl):S3–7. doi:10.1016/S1389-9457(02)00140-5
26. Lanza G, Lanuzza B, Aricò D, et al. Impaired short-term plasticity in restless legs syndrome: a pilot rTMS study. Sleep Med. 2018;46:1–4. doi:10.1016/j.sleep.2018.02.008
27. Lanza G, Lanuzza B, Aricò D, et al. Direct comparison of cortical excitability to transcranial magnetic stimulation in obstructive sleep apnea syndrome and restless legs syndrome. Sleep Med. 2015;16(1):138–142. doi:10.1016/j.sleep.2014.08.016
28. Antelmi E, Mogavero MP, Lanza G, et al. Sensory aspects of restless legs syndrome: clinical, neurophysiological and neuroimaging prospectives. Sleep Med Rev. 2024;76:101949. doi:10.1016/j.smrv.2024.101949
29. Nardone R, Sebastianelli L, Versace V, et al. Contribution of transcranial magnetic stimulation in restless legs syndrome: pathophysiological insights and therapeutical approaches. Sleep Med. 2020;71:124–134. doi:10.1016/j.sleep.2019.12.009
30. Weinstock LB, Walters AS, Paueksakon P. Restless legs syndrome--theoretical roles of inflammatory and immune mechanisms. Sleep Med Rev. 2012;16(4):341–354. doi:10.1016/j.smrv.2011.09.003
31. Geng C, Chen C. Association between inflammatory cytokines and disease severity in restless legs syndrome. Neurol Sci. 2024;45(12):5765–5770. doi:10.1007/s10072-024-07736-w
32. Liu HM, Chu M, Liu CF, Zhang T, Gu P. Analysis of serum vitamin D level and related factors in patients with restless legs syndrome. Front Neurol. 2021;12:782565. doi:10.3389/fneur.2021.782565
33. Mogavero MP, Salemi M, Lanza G, et al. Unveiling the pathophysiology of restless legs syndrome through transcriptome analysis. iScience. 2024;27(4):109568. doi:10.1016/j.isci.2024.109568
34. Benediktsdottir B, Janson C, Lindberg E, et al. Prevalence of restless legs syndrome among adults in Iceland and Sweden: lung function, comorbidity, ferritin, biomarkers and quality of life. Sleep Med. 2010;11(10):1043–1048. doi:10.1016/j.sleep.2010.08.006
35. Akçimen F, Sarayloo F, Liao C, et al. Transcriptome-wide association study for restless legs syndrome identifies new susceptibility genes. Commun Biol. 2020;3(1):373. doi:10.1038/s42003-020-1105-z
36. Allen RP, Picchietti DL, Garcia-Borreguero D, et al. Restless legs syndrome/Willis-Ekbom disease diagnostic criteria: updated International Restless Legs Syndrome Study Group (IRLSSG) consensus criteria--history, rationale, description, and significance. Sleep Med. 2014;15(8):860–873. doi:10.1016/j.sleep.2014.03.025
37. Li YS, Milner PG, Chauhan AK, et al. Cloning and expression of a developmentally regulated protein that induces mitogenic and neurite outgrowth activity. Science. 1990;250(4988):1690–1694. doi:10.1126/science.2270483
38. Uslu FI, Demir E, Güler EM, Koçyiğit A. Circulating levels of cytokines are increased in restless legs syndrome. Sleep Breath. 2021;25(3):1581–1585. doi:10.1007/s11325-020-02218-7
39. Auvinen P, Mäntyselkä P, Koponen H, et al. Elevation of tumor necrosis factor alpha levels is associated with restless legs symptoms in clinically depressed patients. J Psychosom Res. 2018;115:1–5. doi:10.1016/j.jpsychores.2018.09.008
40. Trotti LM, Rye DB, De Staercke C, Hooper WC, Quyyumi A, Bliwise DL. Elevated C-reactive protein is associated with severe periodic leg movements of sleep in patients with restless legs syndrome. Brain Behav Immun. 2012;26(8):1239–1243. doi:10.1016/j.bbi.2012.06.003
41. Sieminski M, Losy J, Partinen M. Restless legs syndrome in multiple sclerosis. Sleep Med Rev. 2015;22:15–22. doi:10.1016/j.smrv.2014.10.002
42. Katz P, Pedro S, Michaud K. Sleep disorders among individuals with rheumatoid arthritis. Arthritis Care Res. 2023;75(6):1250–1260. doi:10.1002/acr.24999
43. Salemi M, Lanza G, Mogavero MP, et al. A transcriptome analysis of mRNAs and long non-coding rnas in patients with Parkinson’s disease. Int J Mol Sci. 2022;23(3):1535. doi:10.3390/ijms23031535
44. Mills KHG. IL-17 and IL-17-producing cells in protection versus pathology. Nat Rev Immunol. 2023;23(1):38–54. doi:10.1038/s41577-022-00746-9
45. Walters AS, Paueksakon P, Adler CH, et al. Restless legs syndrome shows increased silent postmortem cerebral microvascular disease with gliosis. J Am Heart Assoc. 2021;10(11):e019627. doi:10.1161/JAHA.120.019627
46. Manconi M, Garcia-Borreguero D, Schormair B, et al. Restless legs syndrome. Nat Rev Dis Primers. 2021;7(1):80. doi:10.1038/s41572-021-00311-z
47. Laumet G, Bavencoffe A, Edralin JD, et al. Interleukin-10 resolves pain hypersensitivity induced by cisplatin by reversing sensory neuron hyperexcitability. Pain. 2020;161(10):2344–2352. doi:10.1097/j.pain.0000000000001921
48. Guo Z, Zhang J, Liu X, Unsinger J, Hotchkiss RS, Cao YQ. Low-dose interleukin-2 reverses chronic migraine-related sensitizations through peripheral interleukin-10 and transforming growth factor beta-1 signaling. Neurobiol Pain. 2022;12:100096. doi:10.1016/j.ynpai.2022.100096
49. de Souza S, Rosario Claudio J, Sim J, et al. Interleukin-10 signaling in somatosensory neurons controls CCL2 release and inflammatory response. Brain Behav Immun. 2024;116:193–202. doi:10.1016/j.bbi.2023.12.013
50. Hanč P, Gonzalez RJ, Mazo IB, et al. Multimodal control of dendritic cell functions by nociceptors. Science. 2023;379(6639):eabm5658. doi:10.1126/science.abm5658
51. Connor JR, Wang XS, Neely EB, Ponnuru P, Morita H, Beard J. Comparative study of the influence of Thy1 deficiency and dietary iron deficiency on dopaminergic profiles in the mouse striatum. J Neurosci Res. 2008;86(14):3194–3202. doi:10.1002/jnr.21758
52. Zhong X, Drgonova J, Li CY, Uhl GR. Human cell adhesion molecules: annotated functional subtypes and overrepresentation of addiction-associated genes. Ann N Y Acad Sci. 2015;1349(1):83–95. doi:10.1111/nyas.12776
53. Kim MK, Cho YW, Shin WC, et al. Association of restless legs syndrome variants in Korean patients with restless legs syndrome. Sleep. 2013;36(12):1787–1791. doi:10.5665/sleep.3200
54. Moore H, Winkelmann J, Lin L, Finn L, Peppard P, Mignot E. Periodic leg movements during sleep are associated with polymorphisms in BTBD9, TOX3/BC034767, MEIS1, MAP2K5/SKOR1, and PTPRD. Sleep. 2014;37(9):1535–1542. doi:10.5665/sleep.4006
55. Yang Q, Li L, Yang R, et al. Family-based and population-based association studies validate PTPRD as a risk factor for restless legs syndrome. Mov Disord. 2011;26(3):516–519. doi:10.1002/mds.23459
56. Drgonova J, Walther D, Wang KJ, et al. Mouse model for Protein Tyrosine Phosphatase D (PTPRD) associations with restless leg syndrome or Willis-Ekbom disease and addiction: reduced expression alters locomotion, sleep behaviors and cocaine-conditioned place preference. Mol Med. 2015;21(1):717–725. doi:10.2119/molmed.2015.00017
57. Hawrylycz MJ, Lein ES, Guillozet-Bongaarts AL, et al. An anatomically comprehensive atlas of the adult human brain transcriptome. Nature. 2012;489(7416):391–399. doi:10.1038/nature11405
58. Bonnen PE, Wang PJ, Kimmel M, Chakraborty R, Nelson DL. Haplotype and linkage disequilibrium architecture for human cancer-associated genes. Genome Res. 2002;12(12):1846–1853. doi:10.1101/gr.483802
59. Goto-Ito S, Yamagata A, Sato Y, et al. Structural basis of trans-synaptic interactions between PTPδ and SALMs for inducing synapse formation. Nat Commun. 2018;9(1):269. doi:10.1038/s41467-017-02417-z
60. Kwon SK, Woo J, Kim SY, Kim H, Kim E. Trans-synaptic adhesions between netrin-G ligand-3 (NGL-3) and receptor tyrosine phosphatases LAR, protein-tyrosine phosphatase delta (PTPdelta), and PTPsigma via specific domains regulate excitatory synapse formation. J Biol Chem. 2010;285(18):13966–13978. doi:10.1074/jbc.M109.061127
61. Li Y, Zhang P, Choi TY, et al. Splicing-dependent trans-synaptic SALM3-LAR-RPTP interactions regulate excitatory synapse development and locomotion. Cell Rep. 2015;12(10):1618–1630. doi:10.1016/j.celrep.2015.08.002
62. Valnegri P, Montrasio C, Brambilla D, Ko J, Passafaro M, Sala C. The X-linked intellectual disability protein IL1RAPL1 regulates excitatory synapse formation by binding PTPδ and RhoGAP2. Hum Mol Genet. 2011;20(24):4797–4809. doi:10.1093/hmg/ddr418
63. Yoshida T, Shiroshima T, Lee SJ, et al. Interleukin-1 receptor accessory protein organizes neuronal synaptogenesis as a cell adhesion molecule. J Neurosci. 2012;32(8):2588–2600. doi:10.1523/JNEUROSCI.4637-11.2012
64. Yoshida T, Yamagata A, Imai A, et al. Canonical versus non-canonical transsynaptic signaling of neuroligin 3 tunes development of sociality in mice. Nat Commun. 2021;12(1):1848. doi:10.1038/s41467-021-22059-6
65. Yoshida T, Yasumura M, Uemura T, et al. IL-1 receptor accessory protein-like 1 associated with mental retardation and autism mediates synapse formation by trans-synaptic interaction with protein tyrosine phosphatase δ. J Neurosci. 2011;31(38):13485–13499. doi:10.1523/JNEUROSCI.2136-11.2011
66. Tomita H, Cornejo F, Aranda-Pino B, et al. The protein tyrosine phosphatase receptor delta regulates developmental neurogenesis. Cell Rep. 2020;30(1):215–228.e215. doi:10.1016/j.celrep.2019.11.033
67. Wang J, Bixby JL. Receptor tyrosine phosphatase-delta is a homophilic, neurite-promoting cell adhesion molecular for CNS neurons. Mol Cell Neurosci. 1999;14(4–5):370–384. doi:10.1006/mcne.1999.0789
68. Earley CJ, Uhl GR, Clemens S, Ferré S. Connectome and molecular pharmacological differences in the dopaminergic system in restless legs syndrome (RLS): plastic changes and neuroadaptations that may contribute to augmentation. Sleep Med. 2017;31:71–77. doi:10.1016/j.sleep.2016.06.003
69. Bradshaw AD. Diverse biological functions of the SPARC family of proteins. Int J Biochem Cell Biol. 2012;44(3):480–488. doi:10.1016/j.biocel.2011.12.021
70. Blakely PK, Hussain S, Carlin LE, Irani DN. Astrocyte matricellular proteins that control excitatory synaptogenesis are regulated by inflammatory cytokines and correlate with paralysis severity during experimental autoimmune encephalomyelitis. Front Neurosci. 2015;9:344. doi:10.3389/fnins.2015.00344
71. Jones EV, Bouvier DS. Astrocyte-secreted matricellular proteins in CNS remodelling during development and disease. Neural Plast. 2014;2014:321209. doi:10.1155/2014/321209
72. Lively S, Brown IR. Analysis of the extracellular matrix protein SC1 during reactive gliosis in the rat lithium-pilocarpine seizure model. Brain Res. 2007;1163:1–9. doi:10.1016/j.brainres.2007.05.052
73. Kapur N, Friedman R. Oral ketamine: a promising treatment for restless legs syndrome. Anesth Analg. 2002;94(6):1558–1559, table of contents. doi:10.1213/00000539-200206000-00034
74. Silver N, Allen RP, Senerth J, Earley CJ. A 10-year, longitudinal assessment of dopamine agonists and methadone in the treatment of restless legs syndrome. Sleep Med. 2011;12(5):440–444. doi:10.1016/j.sleep.2010.11.002
75. Yepes G, Guitart X, Rea W, et al. Targeting hypersensitive corticostriatal terminals in restless legs syndrome. Ann Neurol. 2017;82(6):951–960. doi:10.1002/ana.25104
76. Ferré S, García-Borreguero D, Allen RP, Earley CJ. New insights into the neurobiology of restless legs syndrome. Neuroscientist. 2019;25(2):113–125. doi:10.1177/1073858418791763
77. Costigan M, Scholz J, Woolf CJ. Neuropathic pain: a maladaptive response of the nervous system to damage. Annu Rev Neurosci. 2009;32:1–32. doi:10.1146/annurev.neuro.051508.135531
78. Woolf CJ, Salter MW. Neuronal plasticity: increasing the gain in pain. Science. 2000;288(5472):1765–1769. doi:10.1126/science.288.5472.1765
79. Hashimoto N, Sato T, Yajima T, et al. SPARCL1-containing neurons in the human brainstem and sensory ganglion. Somatosens Mot Res. 2016;33(2):112–117. doi:10.1080/08990220.2016.1197115
80. Zhu XY, Wu TT, Wang HM, et al. Correlates of nonanemic iron deficiency in restless legs syndrome. Front Neurol. 2020;11:298. doi:10.3389/fneur.2020.00298
81. Walters AS, Rye DB. Review of the relationship of restless legs syndrome and periodic limb movements in sleep to hypertension, heart disease, and stroke. Sleep. 2009;32(5):589–597. doi:10.1093/sleep/32.5.589
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Expression Profile of Pyroptosis-Related Genes and the Associated Regulatory Axis in Primary Gout Patients
Niu S, Lei TY, Dai F, Xie H, Yu X, Zhang Q, Qing Y
Journal of Inflammation Research 2023, 16:1711-1725
Published Date: 19 April 2023
Immune-Related Genes and Immune Cell Infiltration Characterize the Maturation Status of Arteriovenous Fistulas: An Integrative Bioinformatics Study and Experimental Validation Based on Transcriptome Sequencing
Lu P, Wang T, Wan Z, Wang M, Zhou Y, He Z, Liao S, Liu H, Shu C
Journal of Inflammation Research 2024, 17:137-152
Published Date: 8 January 2024
Network Analysis of Osteoarthritis Progression Using a Steiner Minimal Tree Algorithm
Xie Y, Shao F, Ji Y, Feng D, Wang L, Huang Z, Wu S, Sun F, Jiang H, Miyamoto A, Wang H, Zhang C
Journal of Inflammation Research 2024, 17:3201-3209
Published Date: 18 May 2024
Single-Cell Sequencing Combined with Transcriptome Sequencing to Explore the Molecular Mechanisms Related to Psoriasis
E C, Wang R, Meng Z, Zou Y
Clinical, Cosmetic and Investigational Dermatology 2024, 17:2197-2213
Published Date: 3 October 2024